解糖系:
1 分子のグルコースを 2 分子のピルビン酸に分解する経路
UBC/carbohydrate/glycolysis
このページの最終更新日: 2026/04/09広告
概要: 解糖系の全体像
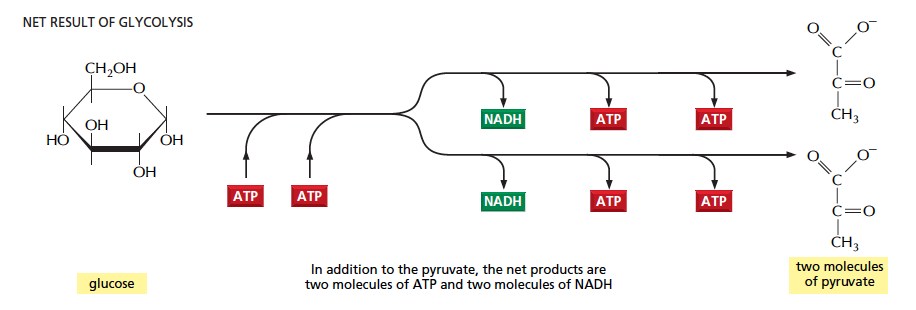
解糖系とは、グルコース が 2 分子の ATP を生み出しつつ各種酵素で分解され、2 分子の ピルビン酸 および 2 分子の NADH を生じる一連の反応である (1)。
"Glycolysis is the sequence of reactions that metabolized one molecule of glucose to two molecules of pyruvate with the concomitant production of two molecules of ATP" (1).
解糖系の全体の反応 net reaction は
Glucose + 2 Pi + 2 ADP + 2 NAD+ →
2 pyruvate + 2 ATP + 2 NADH + 2 H+ + 2 H2O
である (1)。
解糖系の生化学的に重要なポイントは以下の通り。
- 投資・分裂・回収の 3 ステップに分けられる。最終産物のピルビン酸、ATP、NADH はどれも重要。
- 解糖系は、グルコースの分解に 酸素 を必要としない。これは、解糖系が大気中の酸素濃度が増える前に生まれた経路だからと考えられる (1)。つまり解糖系の進化的な起源は非常に古い。
- 解糖系を逆に辿ると 糖新生 に近いが、いくつかの高エネルギー反応の部分で迂回経路を通っている (1)。
- 解糖と糖新生は、同じ細胞内で同時には起こらない (1)。
- 解糖系の中間代謝産物の濃度は、非常に低い (0.1 mmol/kg 以下)。つまり各反応が早い (3)。
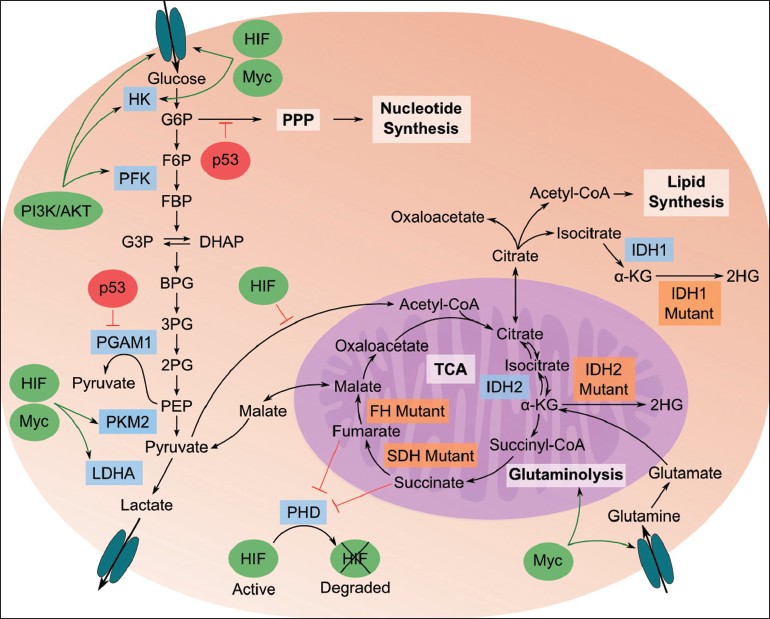
以下の図は、解糖系とピルピン酸の代謝経路などを示したものである (2)。ピルビン酸は、
- 好気的条件下では、アセチル CoA を経て ミトコンドリア の TCA 回路 に入る。
- 嫌気的条件下では、細胞質で 乳酸 lactate に変換される (乳酸発酵)。
- 酵母をはじめとしたいくつかの生物では、細胞質で エタノール に変換される (アルコール発酵)。
広告
「あとがき」で当サイトを参考にしたと書いてくれているラノベです。Kindle Unlimited で読めました。ストーリーと文章が良く、面白かったです。
解糖系に関わる分子の一覧
酵素の一覧
- ヘキソキナーゼ Hexokinase、グルコキナーゼ glucokinase
- ホスホグルコースイソメラーゼ Phosphoglucoseisomerase
- ホスホフルクトキナーゼ Phosphofructokinase
- アルドラーゼ Aldolase
- トリオースリン酸イソメラーゼ Triose phosphate isomerase
- グリセルアルデヒド3-リン酸デヒドロゲナーゼ, Glyceraldehyde 3-phosphate dehydrogenase, GAPDH, G3PDH
- ホスホグリセリン酸キナーゼ Phosphoglycerate kinase, PGK
- ホスホグリセリン酸ムターゼ Phosphoglycerate mutase, PGM
- エノラーゼ Enolase
- ピルビン酸キナーゼ Pyruvate kinase
分解される分子の一覧
グルコース以外の 糖 も、解糖系に合流して分解される。
フルクトース |
肝臓とその他の組織で、異なる経路で解糖系に入る (1)。 肝臓では、フルクトースは fructokinase によって F1P にリン酸化され、glyceraldehyde および DHAP に開裂する。DHAP は解糖系の中間体であり、このままステップ 5 に合流する。Glyceraldehyde は triose kinase によって 3 位でリン酸化され、GAP として解糖系に入る。この経路は 肝臓以外ではヘキソキナーゼ hexokinase が フルクトースを F6P にリン酸化し、ステップ 3 から解糖系に合流する。しかし hexokinase との親和性はグルコースの方が高いため、この経路の効率は低い。 |
ガラクトース |
ガラクトース-1-リン酸 → グルコース-1-リン酸 → グルコース-6-リン酸と変化して解糖系に入る (1)。 |
| マンノース | Hexokinase が マンノース-6-リン酸にリン酸化し、これが phosphomannnose isomerase によって G6P になり解糖系に入る。 |
| グリセロール | 脂肪分解で生じた脂肪酸は β 酸化 に回されるが、グリセロールは glucerol kinase および glycerol phosphate dehydrogenase によって DHAP となる。 ここから解糖によってピルビン酸になる場合と、糖新生によってグルコースまたはグリコーゲンになる場合がある。逆に、グリセロール合成経路は DHAP から分岐する。 |
解糖系からの流出、解糖系への流入
グルコース-6-リン酸 (G6P) |
第一の反応で生じる G6P は代謝の重要な分岐点である。もちろんそのまま解糖系で代謝される場合もあるが、以下の 3 つの重要な流出、流入経路がある。 グリコーゲン合成 (流出) G6P は通常 F6P へと代謝されるが、リン酸基が PGM によって 1 位に転移すると G1P ができる。G1P は UDP との反応を経て グリコーゲン 鎖に付加される。 グルコース合成 (糖新生、流入 & 逆流) 逆に、グリコーゲンから作られた G6P は酵素 glucose-6-phosphatase によってグルコースになる。この反応はほぼ肝臓のみで起こり、他の組織にグルコースを供給するという意義がある。 G6P が G6PD によって脱水素されると、NADP+ が NADPH に還元されるとともに、6-phosphoglucono-δ-lactone ができる。炭素骨格がペントースリン酸回路へ流れる経路である。 |
ジヒドロキシアセトンリン酸 (DHAP) |
DHAP はグリセロール合成の原料となり、最終的に貯蔵脂質である トリアシルグリセロール に取り込まれる。 |
各反応の解説
ここからは、解糖の各反応を詳細にみていく。ポイントは
- 炭素が 6 個ある (C6 と書く) グルコースが、2 個の C3 分子であるピルビン酸になること。
- ATP が生じる反応が 2 つあること。
などの点である。反応を示す図には、炭素番号も記載している。全体像を示す図は 細胞の分子生物学 (Amazon) から引用した。
1. 解糖の開始
トランスポーター GLUT によって細胞内に取り込まれたグルコースは、ヘキソキナーゼ hexokinase によって 6 位でリン酸化され G6P となる。このリン酸基の付加には、以下のような意義がある。
- このリン酸化によってグルコースに大きな負電荷が付与される。これはグルコースが細胞外へ流出するのを防止する効果がある (8)。
- 細胞内のフリーグルコース濃度が下がるため、GLUTを介したグルコースの取り込みが促進される。
- ATP のエネルギーをグルコースに付与している、つまり G6P のエネルギー準位はグルコースよりも高く、その後の代謝反応を受けやすくなる。
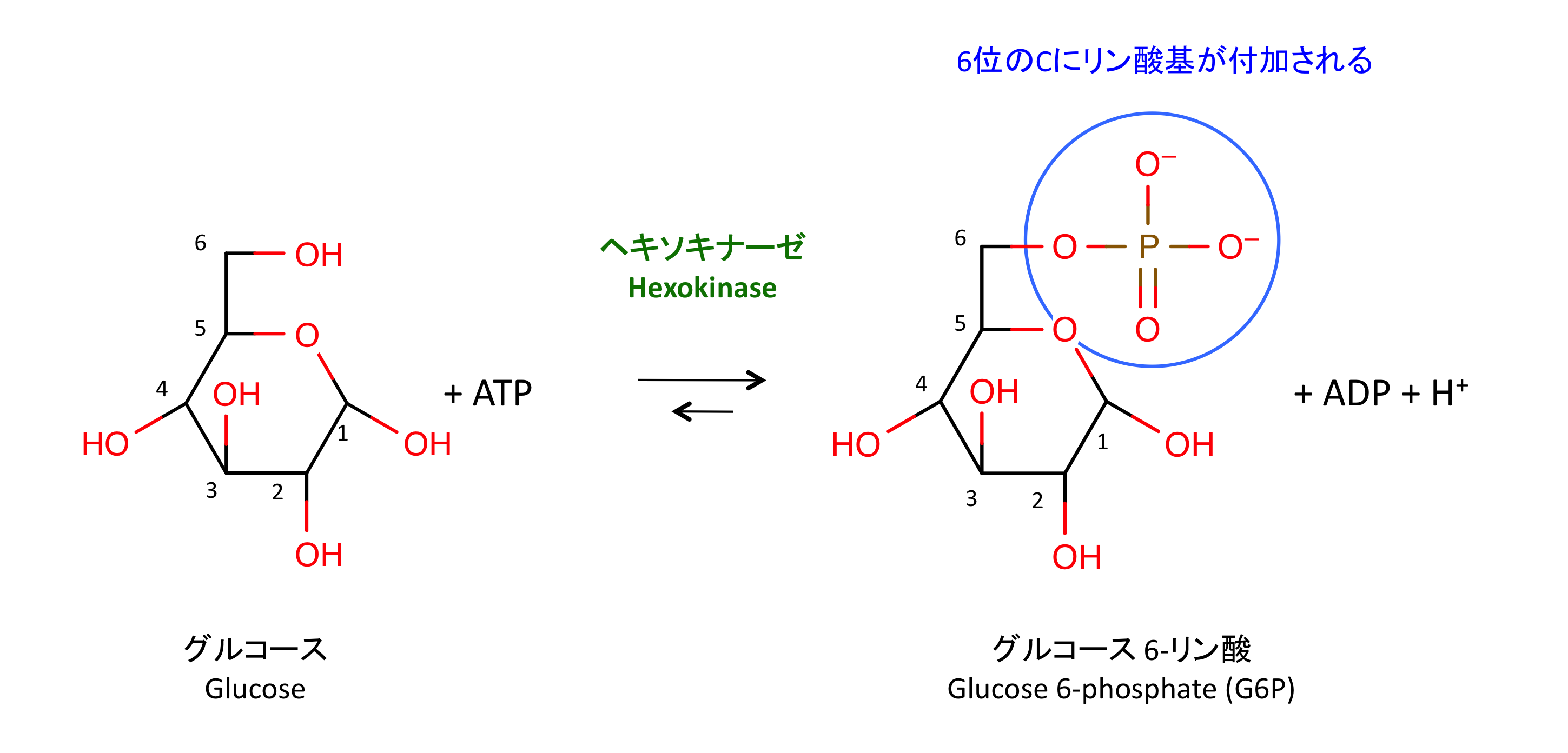
- この反応は不可逆的である。HX は PFK, PK とともに解糖系の律速酵素の一つである (1)。
- G6P はグリコーゲン合成への分岐点である。
- G6P はペントースリン酸回路への分岐点である。NADPH が必要な場合 (= NADP+ が過剰な場合) に G6PD が活性化し、ペントースリン酸経路へ多量の G6P が流れる。
2. フルクトースへの変換
G6P が F6P に変換される。原子の組成が同じまま構造が変わる
C6 分子であるグルコースは、反応 4 において 2 分子の C3 分子に開裂することになる。反応 2 および 3 では G6P をなるべく左右対称な形に近づけようとしており、開裂の準備と考えて良いだろう。
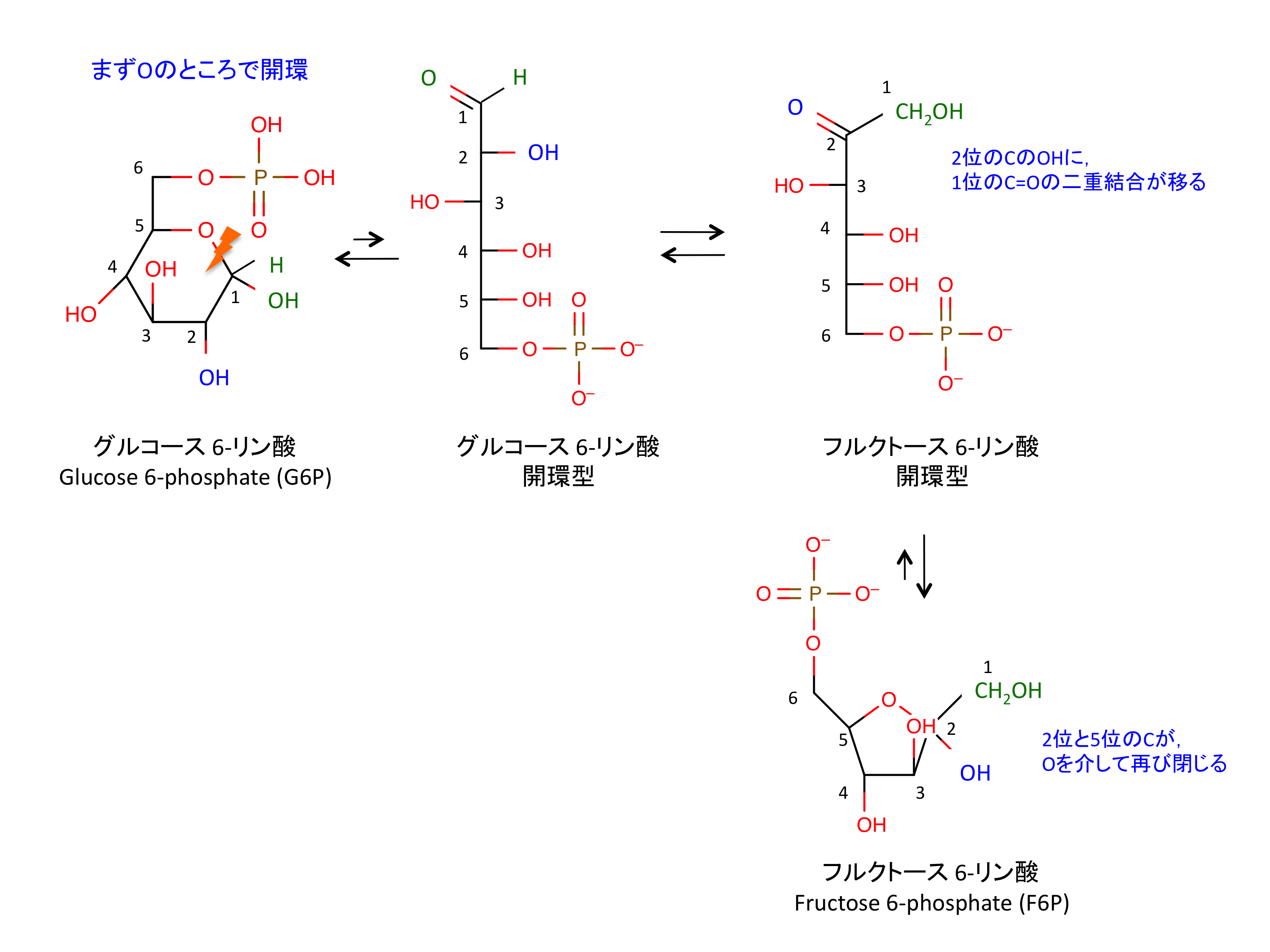
- 肝臓以外の組織では、フルクトースはヘキソキナーゼによって 6 位でリン酸化され、F6P としてここから解糖系に流入してくる (9)。ただし、ヘキソキナーゼはフルクトースよりもグルコースへの親和性が高いので、この経路の効率は低い。
3. F6P のリン酸化
PFK によるこの反応は
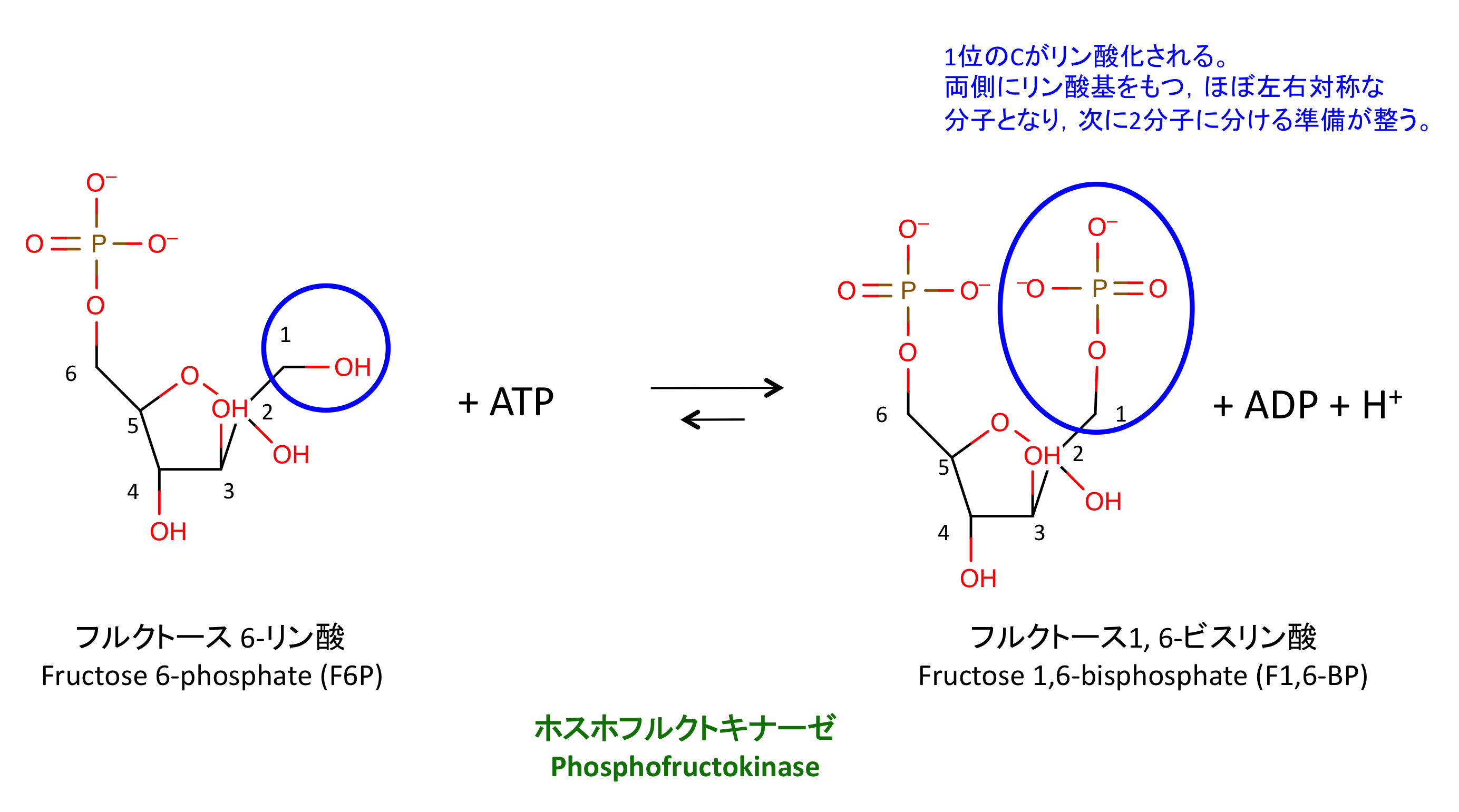
- ヘキソキナーゼ、ホスホフルクトキナーゼ、ピルビン酸キナーゼによる反応が律速段階である。いずれも ATP を消費するリン酸化反応。
4. F1,6P の開裂
ここで 2 つの C3 分子が生じる。この反応は開裂であるが可逆的である。
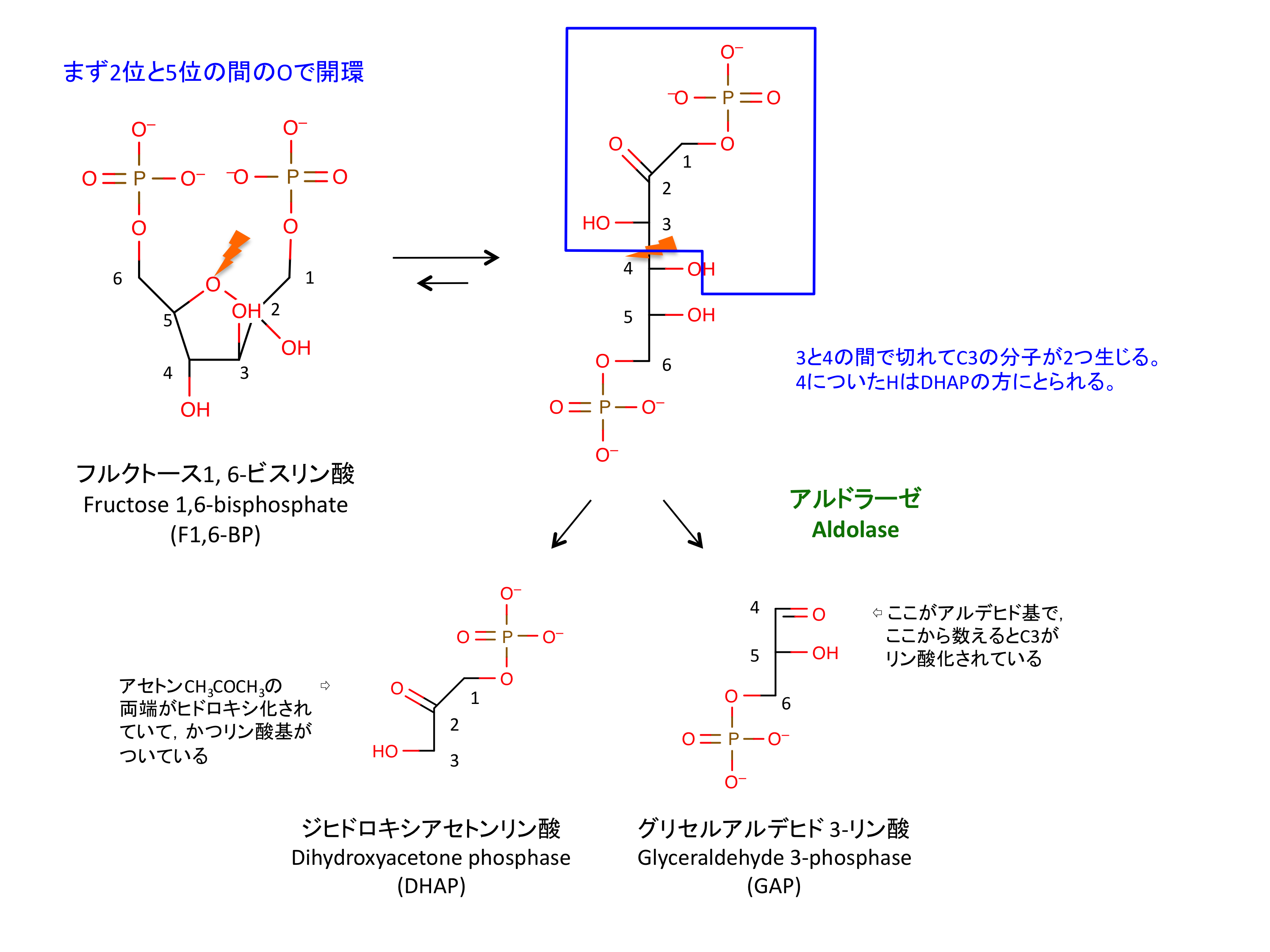
- ここまでの反応で、グルコース 1 分子あたり 2 分子の ATP を消費している。
- GAP はそのまま分解が進むが、DHAP は次の反応で GAP に変換される。
- DHAP はグリセロール合成の原料である (解糖系からの流出)。
5. DHAP の異性化
グルコースはこれによって 2 分子の GAP になる。
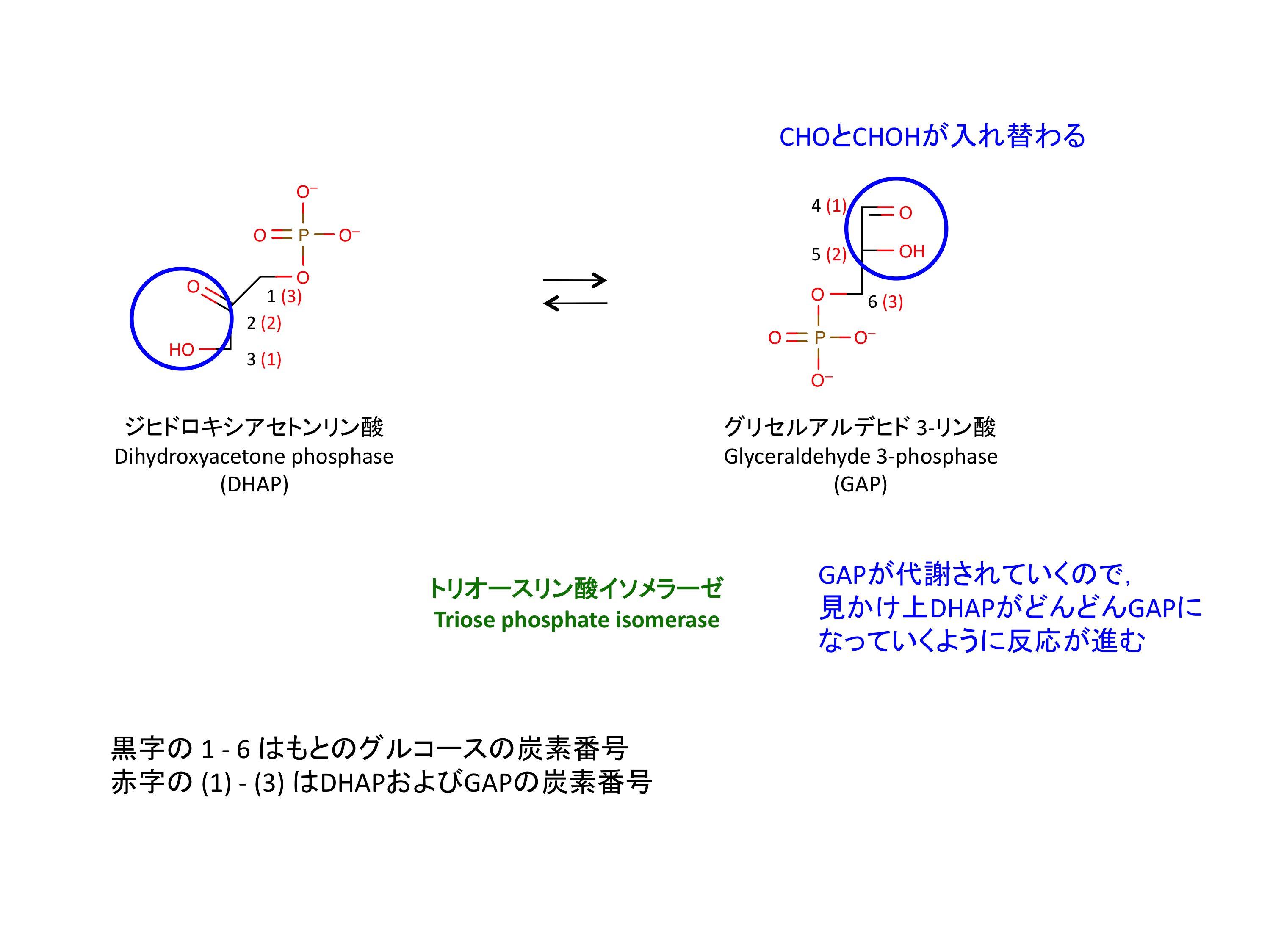
- DHAP と GAP の変換は、可逆的で非常に早い反応である。平衡は DHAP に偏っているが、GAP は反応 6 で取り除かれるので結果的に全体として DHAP が GAP に変わっていく。
- 肝臓では、フルクトースは fructokinase は開裂後に DHAP として解糖系に合流する (1)。Fructokinase によって F1P にリン酸化され、glyceraldehyde および DHAP に開裂する。DHAP はステップ 5 に合流する。Glyceraldehyde は triose kinase によって 3 位でリン酸化され、GAP として解糖系に入る。
6. GAP のリン酸化
この反応は 2 段階。解糖系で唯一の
NAD+ が常に供給されないとこの反応は進まない。嫌気的条件下での乳酸発酵およびアルコール発酵は、NAD+ を再生するための反応である。
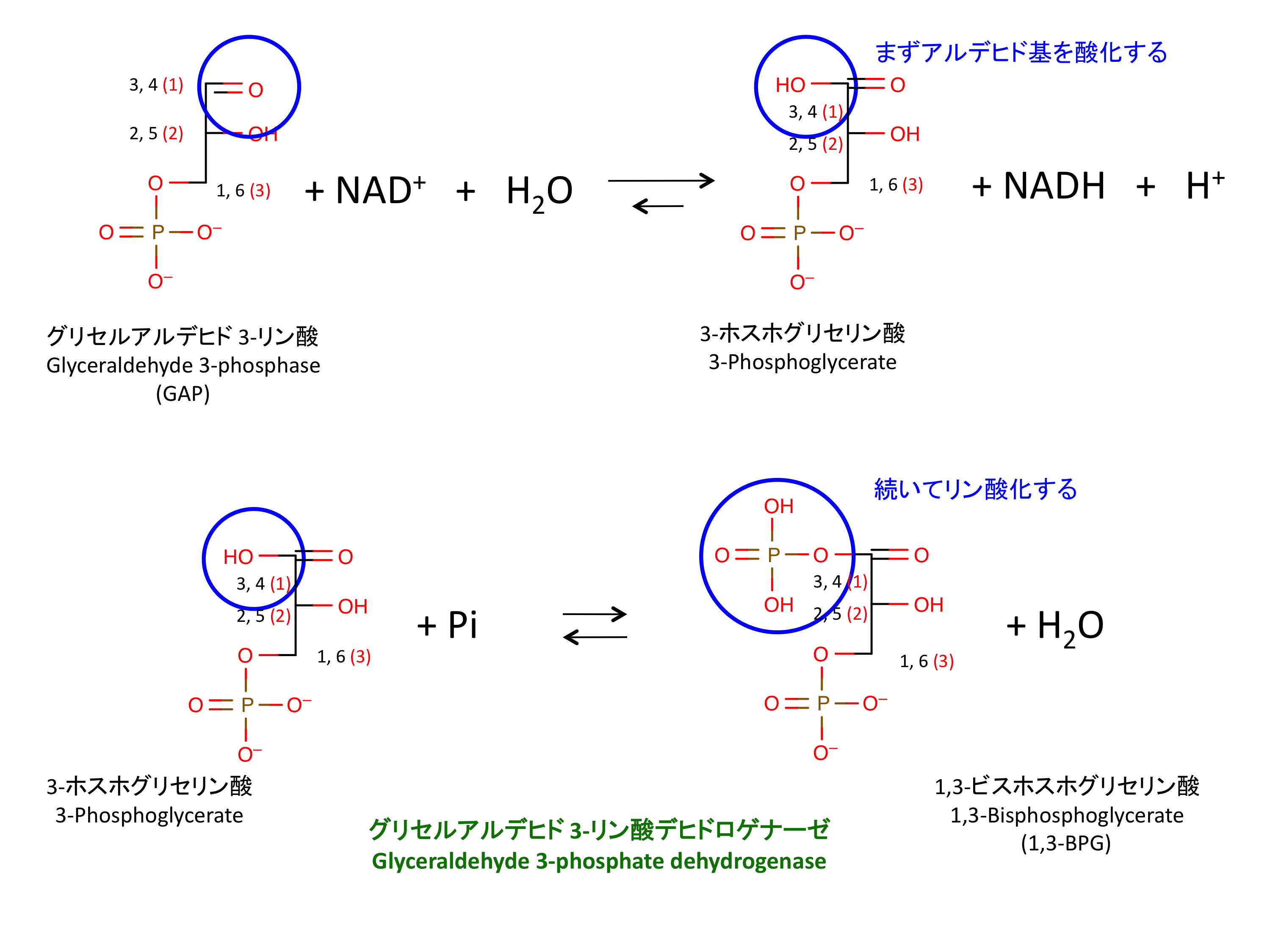
高いリン酸基転移ポテンシャルをもつ 1,3-ビスホスホググリセリン酸 (1,-BPG) が生じる。この分子は次の反応 7 で ADP に Pi を付加し、ATP を産生することができる。
- これはつまり、GAPDH によるリン酸化が分子にかなりのエネルギーを与えているということ。
- 実際、2 番目の反応はかなりの uphill reaction である。
- これは、GAPDH が高エネルギーのチオエステル結合を含む中間体になることによって成される。
この反応を触媒する酵素 GAPDH (G3PDH とも言う) には少なくとも 5 つの分子種がある。上記の反応は、EC.1.2.1.12 G3PDH (phosphorylating) によるものである。
7. 1,3-BPG からの ATP 産生
これは 1,3-BPG が数少ない「ATP よりもエネルギー準位の高い分子」であるからこそ起こりうる反応である。
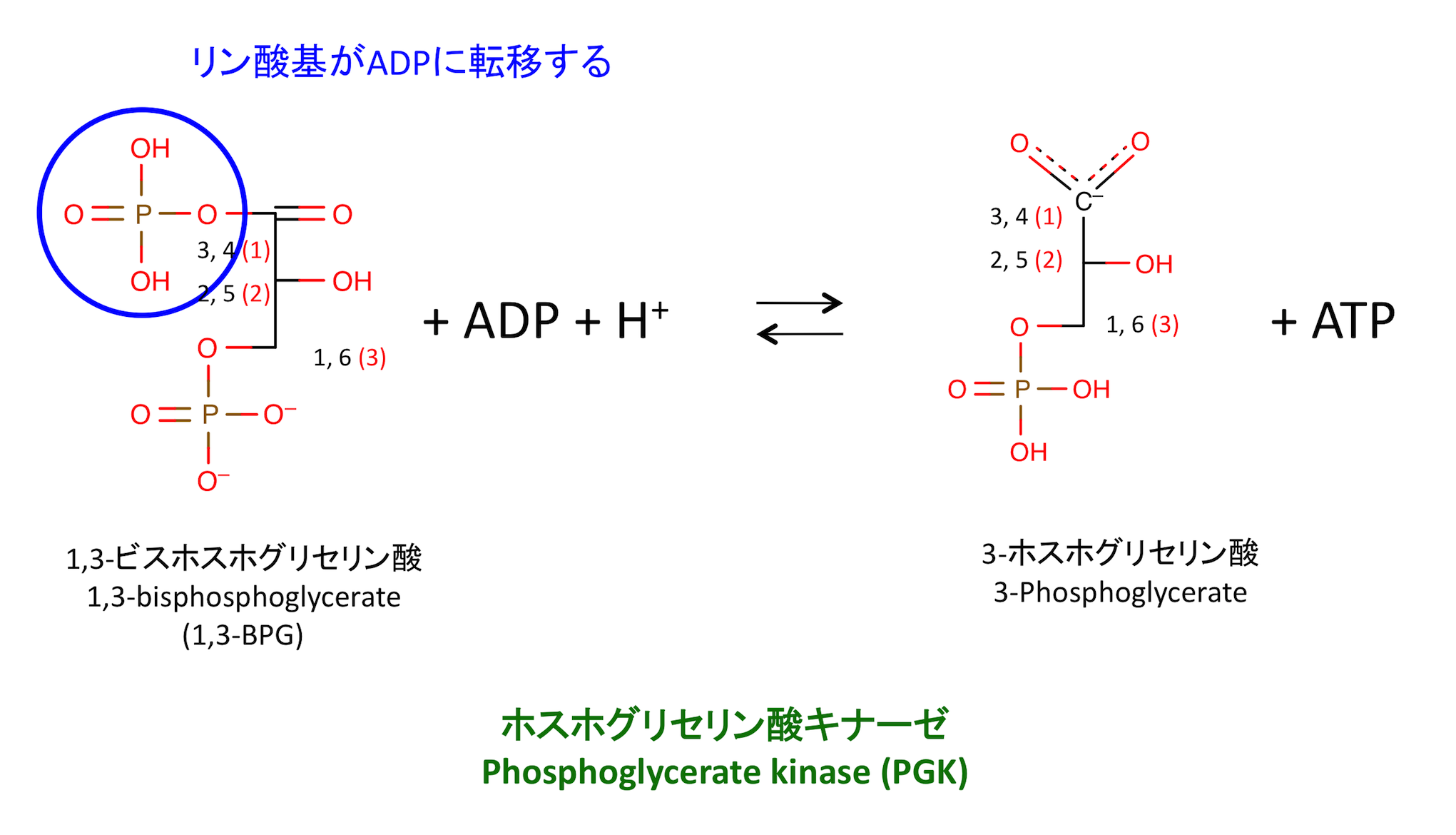
- 解糖系の中間体で ATP よりもATP よりもリン酸基の解離に伴う自由エネルギーが大きいのは、この分子と PEP のみ。「ATP よりもリン酸基の解離に伴う自由エネルギーが大きい中間体が 2 個ある」 = 「解糖系では 2 分子の ATP が作られる」
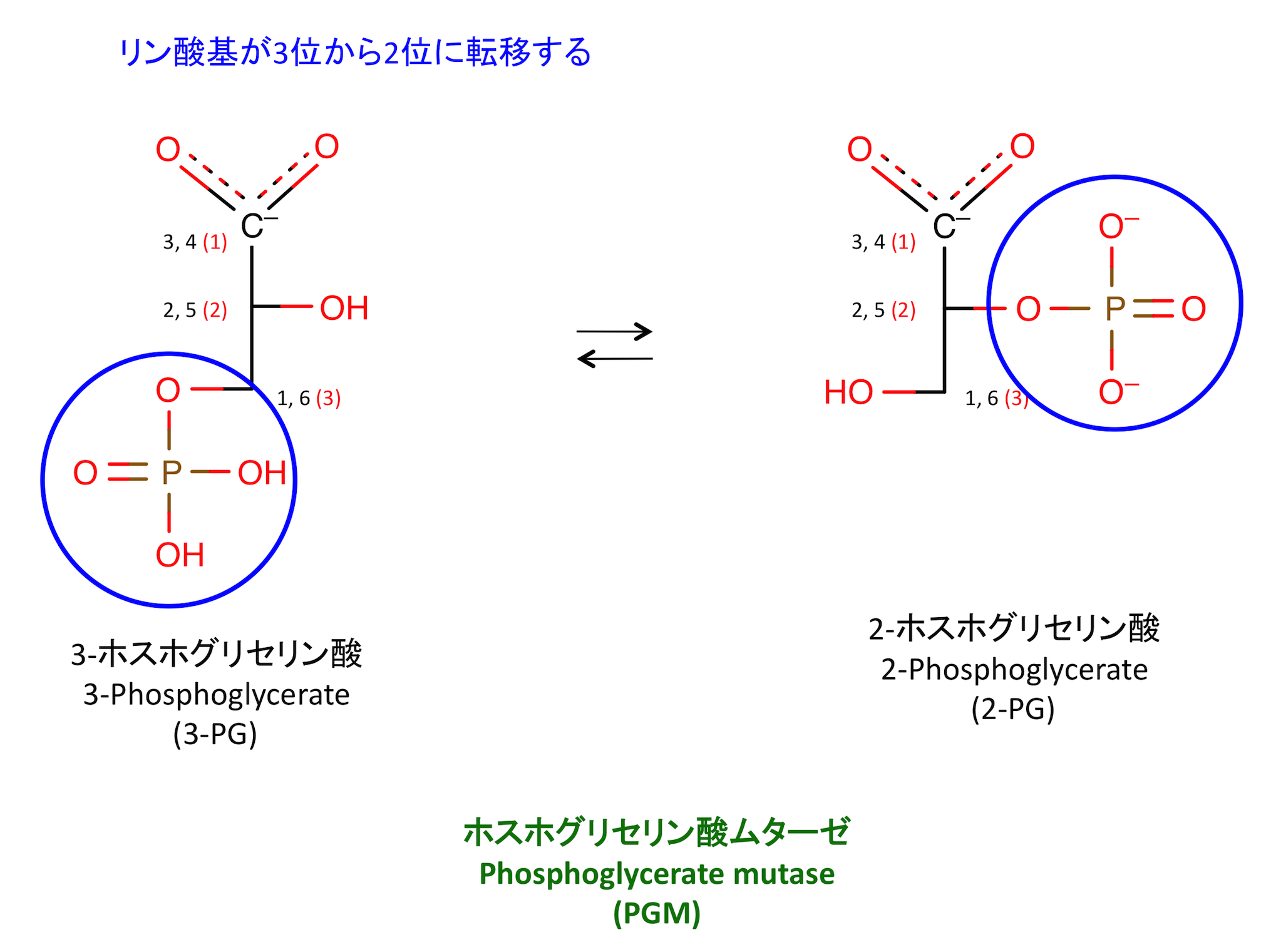
8. リン酸基の分子内転移
Mutase とは、分子内で官能基を移動させる酵素の総称である。
9. 脱水反応
右辺の + H2O は省略 (書き忘れ)。
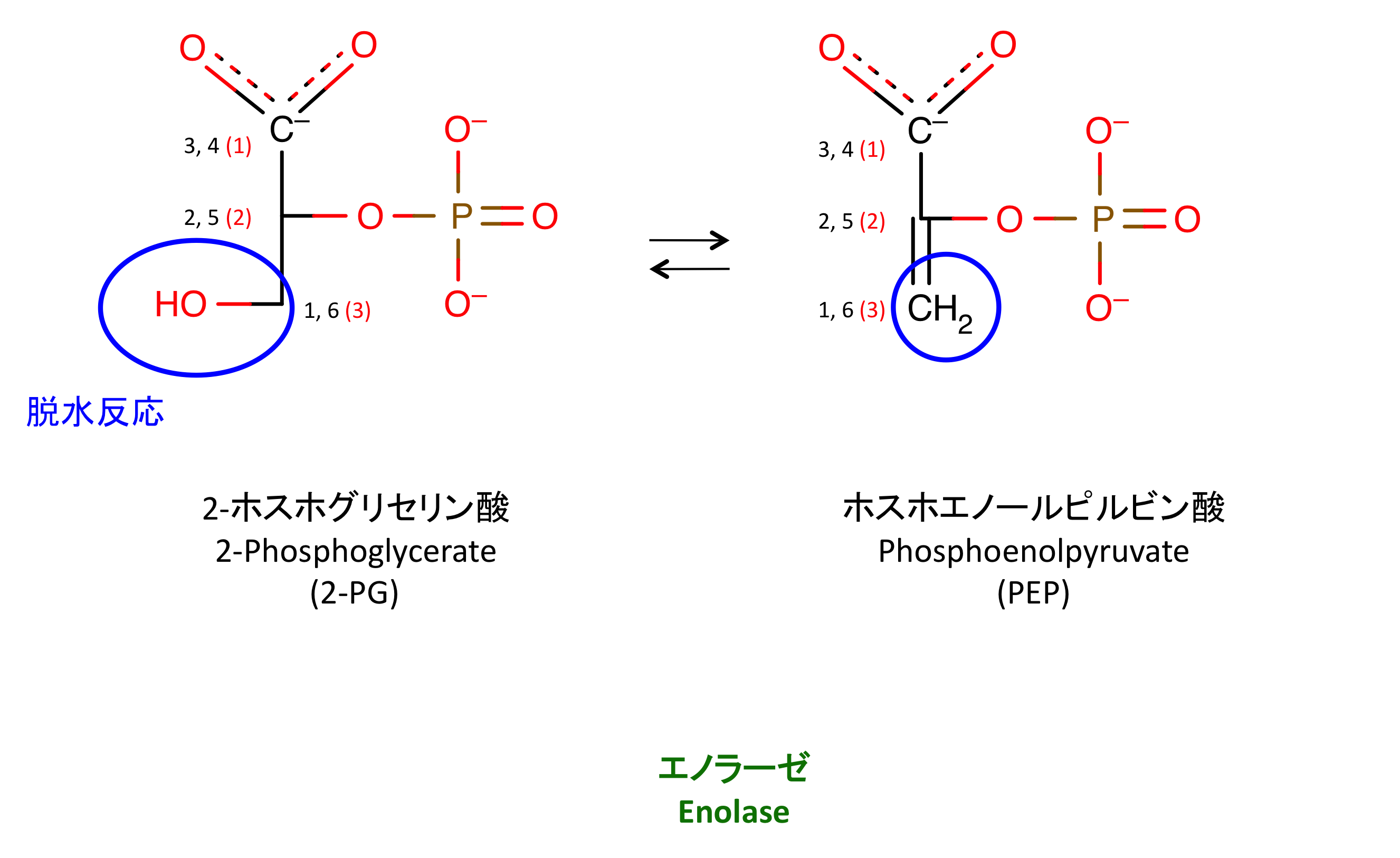
エノール enol は、アルケンの二重結合の片側にヒドロキシ基が置換した アルコール のこと (1)。PEP はエノール型のピルビン酸にリン酸基がついたものである。
通常は、酸素の方が電気陰性度が高いので、エノールの二重結合が O の方へ引っ張られケト型になる。リン酸基があることで不安定な enol 型が保たれ、PEP として存在できる。次の反応でリン酸基が取られると、ピルビン酸はケト型へと変化する。
10. PEP からのリン酸基転移
ホスホエノールピルビン酸から ADP にリン酸基を転移することで ATP を作る。この反応はピルビン酸キナーゼ pyruvate kinase によって触媒される。
この反応は不可逆であり、PK は hexokinase および PFK とともに 3 つの解糖系の律速酵素の一つである
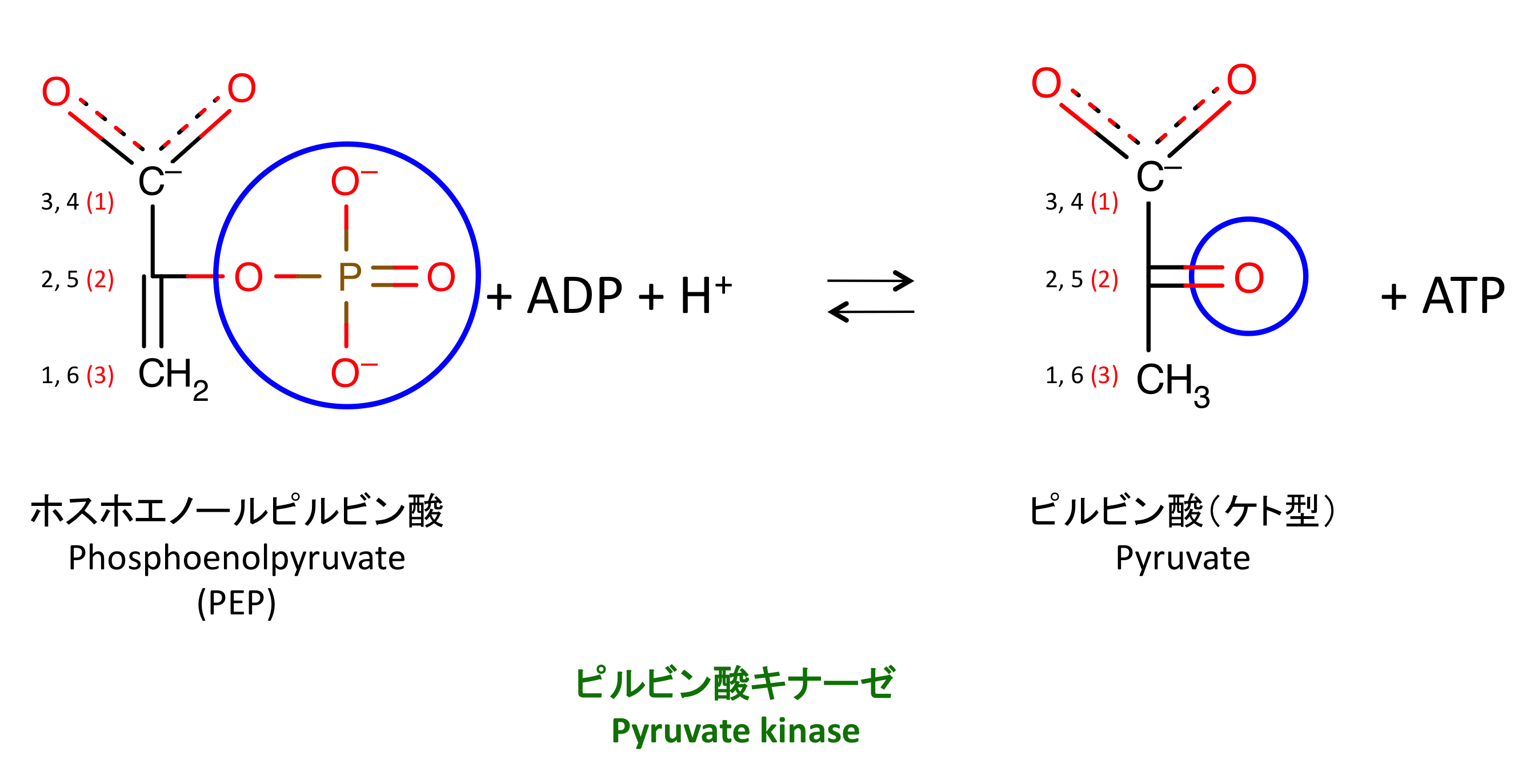
- ピルビン酸 pyruvate は、好気的条件下ではピルビン酸デヒドロゲナーゼ PDH によってアセチル CoA になり、ミトコンドリアにある TCA回路 に入る。
- ピルビン酸は、monocarboxylate transporters, とくに MCT1 によってミトコンドリアまで輸送される (5I)。
- 嫌気的条件化では、乳酸デヒドロゲナーゼ LDH によって乳酸 lactate になる。
解糖系の覚え方
もしもし解糖系
もしもしかめよ〜の音程で歌いましょう。生化学の格言集、語呂合わせ集 も参照のこと。
1.
グルグル 6 リン フル 6 リン フルクの 1, 6 ビスリン酸
アルドで 2 つに脱離して -2 の ATP
2.
次に 1, 3 ビスホスホ +2 の ATP
3 ホス 2 ホスエノラーゼ 血糖値はココ阻害
3.
ホスホのエノールピルビン酸 ココでは後に戻れない
+2 の ATP 作ってなるのがピルビン酸
知識の泉というサイトにあった歌でしたが、残念ながら閉鎖されてしまっているようです。
広告References
Berg et al. 2006a. (Book) . Biochemistry, 6th edition.
|
Berg, Tymoczko, Stryer の編集による生化学の教科書。 巻末の index 以外で約 1000 ページ。 正統派の教科書という感じで、基礎的な知識がややトップダウン的に網羅されている。その反面、個々の現象や分子に対して生理的な意義があまり述べられておらず、構造に偏っていて化学的要素が強い。この点、イラストレイテッド ハーパー・生化学 30版 英語圏ならば学部教育向けにはややレベルが高い印象。しかし、基本を外さずに専門分野以外のことを |
- Shulman 1995a (Review). In vivo regulation of muscle glycogen synthase and the control of glycogen synthesis. Proc Natl Acad Sci USA, 92, 8535-8542.
- de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectrometry of cerebral metabolism. NMR Biomed 16, 339-357.
- Jucker et al. 1997a. 13C and 31P NME studies on the effect of increased plasma free fatty acids on intramusclular glucose metabolism in the awake rat. J Biol Chem 272, 10464-10473.
- Ling et al. 2012a. D-Lactate altered mitochondrial energy production in rat brain and heart but not liver. Nutr Metab 9, 6.
Wei et al. 2009a. Tor1/Sch9-regulated carbon source substitution is as effective as calorie restriction in life span extension. PLoS Genet 5, e1000467.- "TumorMetabolome" by Kathleen A Vermeersch, Mark P Styczynski - DOI: 10.4103/1477-3163.113622; PMID: 23858297. Licensed under CC BY-SA 3.0 via Commons.
- Amazon link: 細胞の分子生物学
: 第 6 版を紹介しています。
Ahern et al. 2017. Biochemistry Free For All.- 解糖系酵素 PDB. Link: Last access 2018/10/25.
{kind=link}
アップデート前、このページには以下のようなコメントを頂いていました。ありがとうございました。
|
2018/10/22 22:26 とてもわかりやすかったです! |
コメント欄
サーバー移転のため、コメント欄は一時閉鎖中です。サイドバーから「管理人への質問」へどうぞ。