嫌気代謝の最終産物 乳酸: 生合成、代謝、構造など
UBC/other_metabolites/organic_acids/lactate
このページの最終更新日: 2026/04/09- 概要: 乳酸とは
- 生合成
- 乳酸を生合成する組織
- 分解
- 血中動態
- シグナル分子としての乳酸
- アルコール発酵
- 乳酸菌とバクテリアの乳酸代謝 → プロバイオティクス へ
広告
概要: 乳酸とは
乳酸 lactic acid は、図のような構造をもつ 解離定数 pKa = 3.86 の有機酸である (14I)。生理的 pH 域では、ほぼ完全に乳酸イオン lactate と H+ に解離している。

L-乳酸 (左) および L-乳酸イオン (右)。
乳酸には光学異性体のL-乳酸とD-乳酸があり、真核生物で主に代謝に利用されるのはL-乳酸である。このサイトでは、とくに断らない限りはL-乳酸について述べる。
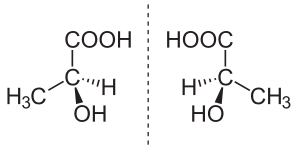
L-乳酸 (左) および D-乳酸 (右)。
ヒト、マウスなど一般的な生化学の教科書に載っているような代謝系では、解糖 によって生じた ピルビン酸 が嫌気的条件下では 乳酸脱水素酵素 (乳酸デヒドロゲナーゼ, lactate dehydrogenase によって L-乳酸 L-lactate に変換される。
このページでは、とくに断りがない限り 哺乳類 の乳酸代謝系について述べ、バクテリア の乳酸代謝については最後の項目にまとめる。
乳酸は、以下のような生化学的に重要な特徴をもっている。
- 嫌気代謝マーカー
- 糖新生 gluconeogenesis の原料
- 脂肪分解の阻害
- 炎症の抑制
広告
乳酸の生合成
乳酸の生合成経路として有名なものは、嫌気的条件下でのピルビン酸からの生合成がある。この反応には、以下の ような意義がある (6D, 8)。
- 嫌気的条件下では酸化的リン酸化の速度が低下し、TCA 回路 の回転速度が遅くなってピルビン酸が過剰になる。このピルビン酸を取り除くという意義 (6D)。
- また、酸化的リン酸化の速度が遅くなると、TCA 回路の生成物の一つである NADH が蓄積する。これは TCA 回路の回転をますます遅くしてしまうので、乳酸合成によって NAD+に再変換する (6D)。また、解糖系に必要な NAD+ を再生産する。
- TCA 回路のない赤血球でも、NAD+ と NADH のバランスは細胞の恒常性を保つために重要である。ピルビン酸を除去して、細胞のレドックス状態を維持するために乳酸を合成する (8)。

ピルビン酸からの生合成は上のように行われる。C=O が H をもらって CH-OH になるので、還元反応である。反応は、乳酸デヒドロゲナーゼ lactate dehydrogenase によって触媒される。
この反応では、最初に酵素と NADH が結合し、乳酸の後に最後に NAD+ が解離する (8)。
グルコースからの乳酸生成、いわゆる「嫌気的条件下での解糖」は、
Glucose + 2 Pi + 2 ADP → 2 Lactate + 2 ATP + 2 H2O
として表すことができる。最終的にグルコース 1 分子から 2 分子の ATP を得ていることになる。
乳酸を生合成する組織
乳酸は、嫌気代謝が活発な組織で合成される。赤血球 erythrocyte はミトコンドリアを持たず、また運動中の筋肉は酸素不足に陥っているので、これらが乳酸生成の主要な器官である (8)。
主に骨格筋、赤血球、脳、腸管で生じるとする論文もある (4I)。脳では、刺激によって神経が興奮した後に局所的に乳酸濃度が上がり、5 秒後には元に戻る (12)。
乳酸の分解
乳酸はグルコース代謝の最終産物であり、さらに代謝されるためにはピルビン酸に戻されなければならない (8)。骨格筋、赤血球などの組織で合成された乳酸は、血液中に放出され、以下の2通りの経路で分解される。
TCA 回路
心筋細胞に代表されるいくつかの細胞では、膜に乳酸およびピルビン酸を取り込むキャリアがある。取り込まれた乳酸はピルビン酸に変換され、アセチル CoA として TCA 回路に入る。
これは、それらの細胞が乳酸をエネルギー源として使っているということであり、血中のグルコースを節約して骨格筋に優先的に使わせるという意義もある。
糖新生
肝臓に取り込まれ、ピルビン酸を経て糖新生によりグルコースになる。この
グルコース → ピルビン酸 → 乳酸 → 肝臓へ移動 → 乳酸 → ピルビン酸 → グルコース
という一連の過程を乳酸回路 lactic acid cycle またはコリ回路 Cori cycle という (8)。
血中動態
乳酸は、血中では一価の陰イオン CH3C(OH)COO- として存在する (4I)。血中乳酸濃度は、体内の酸化還元状態を反映しているとされ、循環不全や運動で高値を示す (4I)。主な発生源は、嫌気代謝に陥りやすい筋肉と、ミトコンドリアをもたない赤血球である。
乳酸は酸であるため、血中乳酸濃度が高くなると pH が酸性に傾くアシドーシス acidosis を引き起こす。
> アシドーシスは、例えば筋細胞内では過剰な H+ がタンパク質の Ca2+ 結合と競合して悪影響を示す (7I)。
- ただし、アシドーシスの影響は、血液の pH のみで決まるわけではない。
- 例えば乳酸よりも CO2の方が細胞膜を通過しやすいので、細胞への影響は大きい。
> 一般に、心筋はアシドーシスによる機能不全を起こしやすい (7I)。
- Ca2+ シグナルが重要だからかもしれない。
- カメの心筋は、アシドーシスや無酸素状態 anoxia への耐性が強いことが知られている。
> 腎不全患者などの透析外液を分析し、血中の代謝産物量を測定した論文 (9R)。
- 乳酸およびピルビン酸量は透析中に上昇する。合成が起こっていることを示唆する。
- ただし、乳酸/ピルビン酸比から考察するに、これは低酸素が原因ではない。補酵素の欠乏かも。
- 腎不全の原因 (糖尿病性であるか否かなど)によって、代謝産物量と透析中の変化が異なる。
- 血中 Ala, Val 濃度は透析中ほぼ一定、クレアチニンは指数関数的に減少した。
広告
「あとがき」で当サイトを参考にしたと書いてくれているラノベです。Kindle Unlimited で読めました。ストーリーと文章が良く、面白かったです。
シグナル分子としての乳酸
乳酸は、受容体 GPR81 を介してシグナル分子としても働く (12)。
> 脂肪細胞では、cAMP と PKA 活性を低下させ、脂肪分解 lipolysis を阻害する方向に働く (12)。
> 活性化される遺伝子として、c-fos, c-jun, c-ets, Hyal-1, Hyal-2, CD44, caveolin-1 がある (12)。
> 乳酸は、HCA1 を介してToll-like receptor の活性化による肝臓、膵臓の炎症を抑制する (14R)。
- マウスのマクロファージに LPS で炎症を誘発すると、培地中の乳酸量が増える。
- 予め培地に 15 mM 乳酸を入れておくと、カスパーゼ1などの炎症マーカーの発現が抑制される。
- NF-κB Ser536 のリン酸化も抑制される。これは、TLR-4 による NF-κB 活性化の指標。
- HCA1 をノックダウンすると、乳酸による炎症抑制がみられなくなる。
- in vivo でも、乳酸は HCA1 依存的に肝臓と膵臓の LPS 依存的な炎症を抑制した。
- ノックダウンの効果など、HCA1 量と表現型に全く相関がないのが気にかかる。
乳酸は、血液脳関門を monocarboxylate transporter (MCT) の作用で通過することができ (1I)、脳には約 0.5 - 1.5 mmol/kg の濃度で存在する (2)。この濃度は MRS では通常見えない (3)が、低酸素状態に曝された動物の脳では顕著なピークとして観察される。
低血糖時に、脳の機能を維持する役割をもつが、これは乳酸がエネルギー源として利用されるためではない (11R)。乳酸は低血糖時にグルコースの代わりのエネルギー源となるが、それはエネルギー全体の約10%に過ぎない (1I)。それよりも、乳酸が受容体 GPR81 を通して作用することにより、グルコースが効率的に代謝されるようになることが重要と考えられている。
アルコール発酵
> キンギョ、Crucian carp など一部の魚類は、乳酸からエタノールを作る (5R)。
- 乳酸による血液 pH の上昇 (アシドーシス acidosis)が起こらないという利点がある。
- Crucian carp では、この反応は筋肉で起こる (10I)。
- このように、発酵に代謝を切り替えられる生物を通性嫌気性生物という。バクテリアが有名である。
広告
References
Boumezbeur et al. 2010a. The contribution of blood lactate to brain energy metabolism in humans measured by dynamic 13C nuclear magnetic resonance spectroscopy. J Neurosci, 30, 13983-13991.de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectroscopy of cerebral metabolism. NMR Biomed 16, 339-357.Govindaraju et al. 2000a. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed 13, 129-153.宮口ほか 2002a. 電気化学検出高速液体クロマトグラフィーによる血中乳酸の検出. 分析化学 51, 703-706.van den Tshillart et al. 1989a. Fish muscle energy metabolism measurement by in vivo 31P-NMR during anoxia and recovery. Am J Physiol 256, R922-R929.deRoos 1994a. Plasma ketone, glucose, lactate, and alanine levels in the vascular supply to and from the brain oh the spiny dogfish shark (Squalus acanthias). J Exp Zool 268, 354-363.Jackson et al. 1991a. 31P-NMR study of normoxic and anoxic perfused turtle heart during graded CO2 and lactic acidosis. Am J Physiol 260, R1130-R1136.Berg ed. 2006a (Book). Biochemistry, chapter 16. W.H. Freeman and Company, NY.安藤 2012a .1H-NMR メタボロミクスの基礎技術開発と透析治療への応用. 東北大学博士論文.Lardon et al. 2005a. 1H-NMR study of the metabolome of an exceptionally anoxia tolerant vertebrate, the crucian carp (Carassius carassius). Metabolomics 9, 311-323.Herzog et al. 2013a. Lactate preserves neuronal metabolism and function following antecedent recurrent hypoglycemia. J Clin Invest 123, 1988-1998.Bergersen & Gjedde 2012a (Review). Is lactate a volume transmitter of metabolic states of the brain? Front Neuroenergetics 4, 5.- ページ編集に伴い削除
Hoque et al. 2014a. Lactate reduces liver and pancreatic injury in Toll-like receptor- and inflammasome-mediated inflammation via GPR81-mediated suppression of innate immunity. Gastroenterology 146, 1763-1774.
コメント欄
サーバー移転のため、コメント欄は一時閉鎖中です。サイドバーから「管理人への質問」へどうぞ。