TCA回路: ほとんどの栄養素が最終的に酸化される分子経路
UBC/aa_carbo_lipid/carbohydrate/tca_cycle
このページの最終更新日: 2026/04/09広告
概要: TCA 回路の全体像
- 反応のほとんどは、ミトコンドリア のマトリックスで行われる。
- 栄養素の多くは アセチル CoA として TCA 回路に入り、最終的に 二酸化炭素 にまで酸化される。
- その過程で、高エネルギー分子 NADH、FADH2 および GTP が作られる。
- アミノ酸代謝、尿素回路、糖新生 など多くの代謝経路の仲立ちをする。
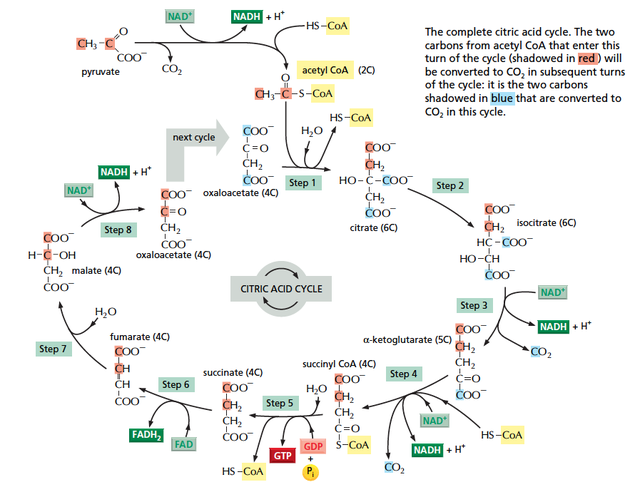
食べ物から取り出したエネルギーを、高エネルギー分子 NADH および FADH2 に変換する経路とも考えられる。また、NADH や FADH2 は電子の運搬体であるため、high energy electron を食べ物から取り出す経路という表現も可能である。
TCA 回路の全体の反応は、以下の式で表すことができる。
アセチル CoA + 3NAD+ + FAD + GDP + Pi + 2H2O
↓
2CO2 + 3NADH + FADH2 + GTP + 2H+ + CoA
TCA 回路はクエン酸回路 citric acid cycle、クレブス回路 Krebs cycle とも呼ばれる。citric の発音は [sitrik] であり、「サイトリック」ではないので注意すること。
広告
「あとがき」で当サイトを参考にしたと書いてくれているラノベです。Kindle Unlimited で読めました。ストーリーと文章が良く、面白かったです。
TCA 回路に関わる分子の一覧
酵素の一覧
- ピルビン酸デヒドロゲナーゼ Pyruvate dehydrogenase
- クエン酸シンターゼ Citrate synthase
- アコニターゼ Aconitase
D-イソクエン酸デヒドロゲナーゼ D-isocitrate dehydrogenase (IDH): 律速酵素- α-ケトグルタル酸デヒドロゲナーゼ α-ketoglutarate dehydrogenase
- サクシニル-CoA シンターゼ Succinyl CoA synthase
- コハク酸デヒドロゲナーゼ Succinate dehydrogenase
- フマラーゼ Fumarase
- リンゴ酸デヒドロゲナーゼ Malate dehydrogenase
分解される分子の一覧
有機酸、アミノ酸など分子の種類に応じて表を分けている。
ピルビン酸 |
ピルビン酸デヒドロゲナーゼ複合体に酸化されてアセチル CoA として TCA 回路に入る (このページの反応 0)。 ピルビン酸カルボキシラーゼに触媒されオキサロ酢酸として入る経路もある。これは補充反応 anaplerosis と呼ばれる。 |
酢酸 |
アセチル CoA リガーゼによって補酵素 A と結合し、アセチル CoA になる。これが TCA 回路で酸化される。 |
|
分岐鎖アミノ酸である Val, Leu, Ile には共通の分解経路が存在する。
|
TCA 回路からの流出
| アセチル CoA |
アセチル CoA が TCA 回路に入らなかった場合、脂肪酸合成 に使われる経路がある。酵素 ACC がアセチル CoA からマロニル CoA を作るのが脂肪酸合成の第一段階である。マロニル CoA には 脂肪酸の β 酸化 を抑制する作用があり、脂肪酸の合成と酸化が同じ細胞で同時に起こらないようになっている。 |
α-KG |
2-oxoglutarate (2-OG) とも呼ばれる。この総説 で詳しく解説されているように、重要な代謝中間体である。 重要経路: α-Ketoglutarate からは グルタミン酸 が合成される。神経系では Glu の一部がさらに GABA に変換され、その後 TCA 回路に戻ってくる経路がある。これは GABA shunt と呼ばれ、TCA 回路のバイパス経路と考えられる。 |
リンゴ酸 |
NADP-linked malic enzyme (EC 1.1.1.40) によって脱炭酸され、ピルビン酸になる反応がある。肝臓、腎臓および脳に存在する (9I)。Pyruvate recycling とよばれる |
オキサロ酢酸 |
オキサロ酢酸は、様々な生合成反応の出発点である。
|
広告
各反応の解説
ここからは、TCA 回路の各反応を詳細にみていく。反応の番号は教科書によって異なっているようであるが、このサイトでは 細胞の分子生物学 (Amazon link) の番号に従っている。
0. ピルビン酸の酸化
ピルビン酸の酸化は、解糖系と TCA サイクルを結びつける重要な反応である。3 つの酵素と 5 つの補酵素を必要とする複雑な反応であるが、全体として以下の式で表すことができる。
Pyruvate + CoA + NAD+ -> Acetyl-CoA + CO2 + NADH + H+
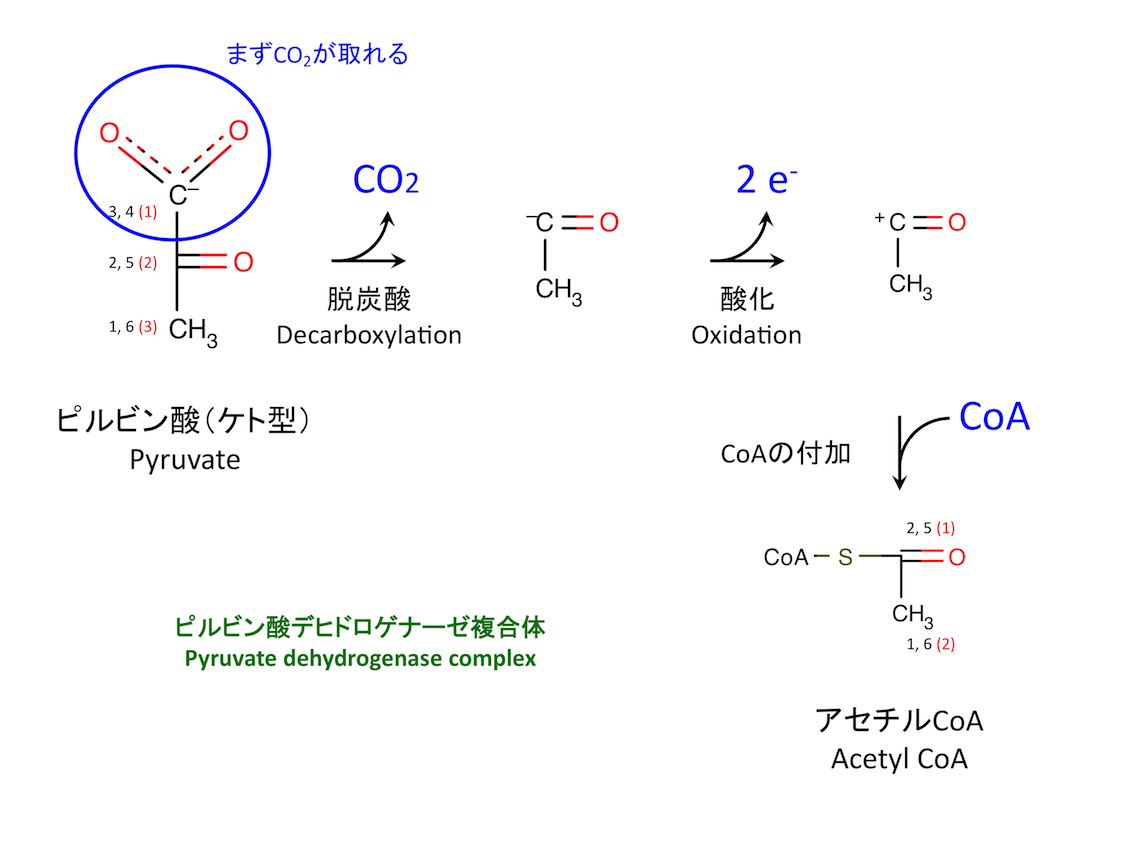
黒字の 1 - 6 は、ピルビン酸が解糖系に由来する場合のグルコースの炭素番号。赤字の 1 - 3 は、ピルビン酸またはアセチル CoA の炭素番号。
解糖系の最終産物であるピルビン酸 pyruvate は、好気的条件下ではミトコンドリアへ輸送されアセチル CoA になる (1)。嫌気的条件化では乳酸 lactate またはエタノール ethanol に代謝される。この分岐は生化学上極めて重要である。
> 脊椎動物の細胞では、ATP の 95% 以上が好気呼吸で産生される (10D)。
- 通常の酸素濃度の場合である。
- グルコースの大部分が水と二酸化炭素になり、
乳酸になるのは 4% 程度 。
> アセチル CoA の合成反応は
- つまり、いったんこの反応が起こったら、グルコースは酸化されるか脂質になるかという運命しかない。
- 脂肪酸合成の経路は acetyl CoA から分岐し、分解された脂肪は acetyl CoA になる。
1. クエン酸の合成
アルドール縮合および脱水反応で、クエン酸シンターゼ citrate synthase に触媒される (1)。
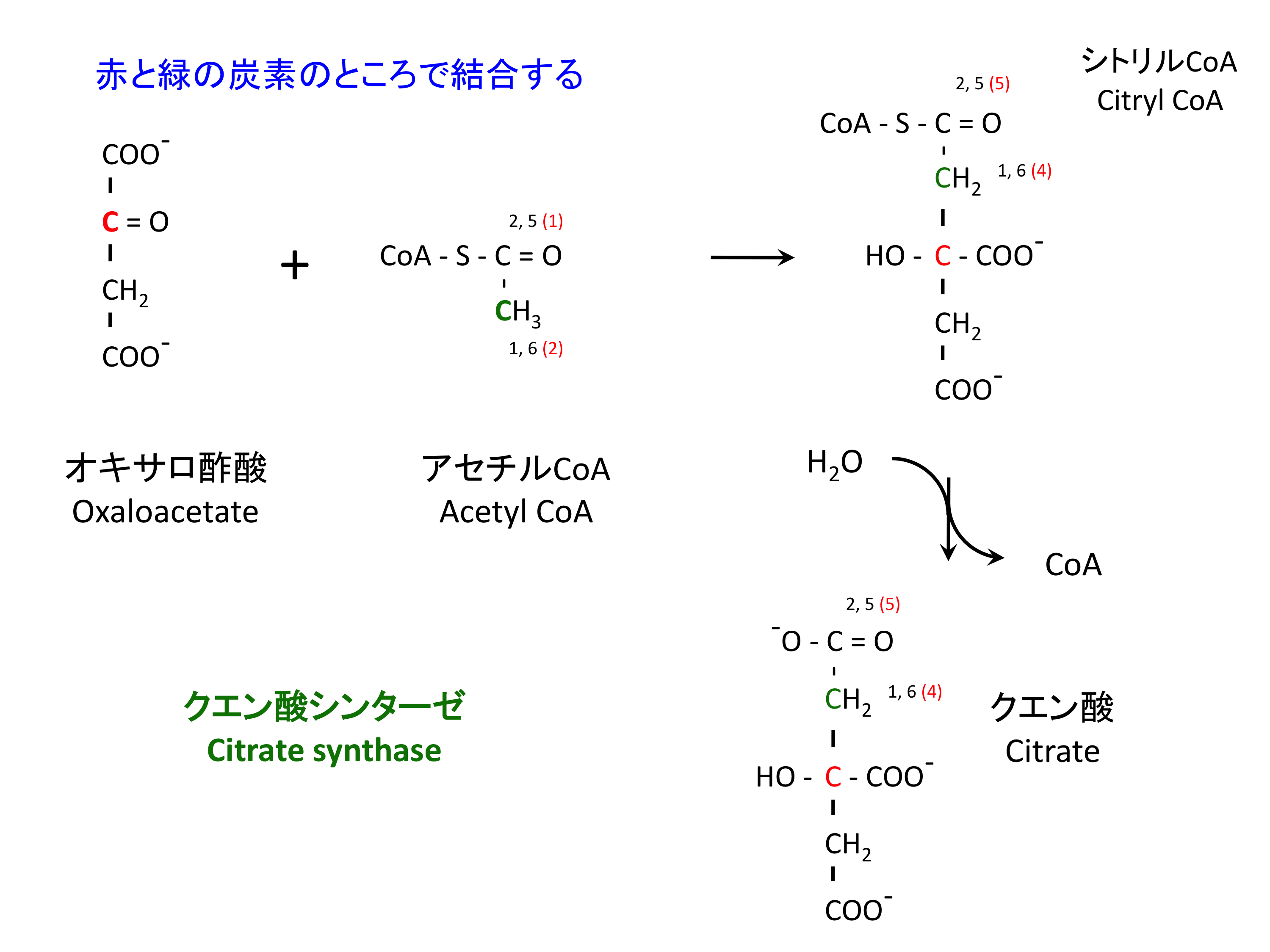
> アセチル CoA の加水分解が間違って起こらないようなメカニズムがある (1)。
- Citrate synthase は最初にオキサロ酢酸に結合しないと、アセチル CoA と結合することができない。
- 加水分解の活性中心は、citryl CoA と酵素の相互作用で正しい位置に来るようになっている。
- 1, 2, 5, 6 は、グルコースが解糖を経て TCA 回路に入ってきた場合の、もとのグルコースの炭素番号。
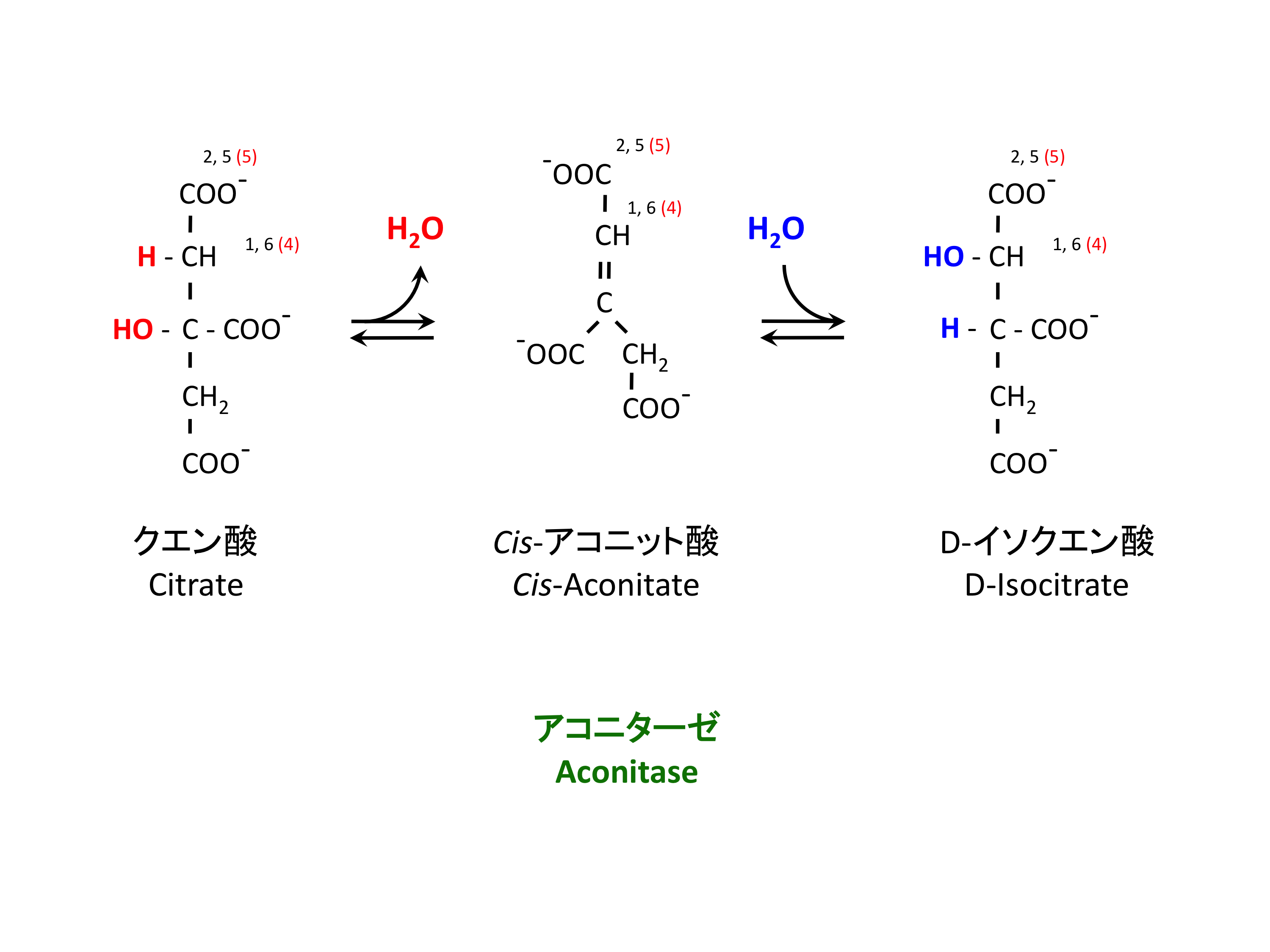
2. クエン酸の異性化
クエン酸は安定すぎるため、今後の反応のために H と OH の位置を入れ替える反応である。実際は脱水反応 dehydration と水和反応 hydration が連続して起こっている。反応を触媒するのはアコニターゼ aconitase である。
> アコニターゼは、細胞質で鉄調節タンパク質としても機能する (→ See blog)。
- ヘム heme を使わずに鉄 iron と結合する珍しいタンパク質である。
3. イソクエン酸の酸化・脱炭酸
ここから 4 つの酸化還元反応が連続することになり、イソクエン酸の酸化はその第一段階である。この反応は
イソクエン酸 + NAD+ -> α-ケトグルタル酸 + CO2 + NADH
と表すことができる。NADH を生成する最初の反応である。
酵素 D-イソクエン酸デヒドロゲナーゼが行うのは、水素を奪う反応である。中間体として生じるオキサロコハク酸は青字の COO- 部分が不安定であり、酵素に結合したままの状態で二酸化炭素を生じる。
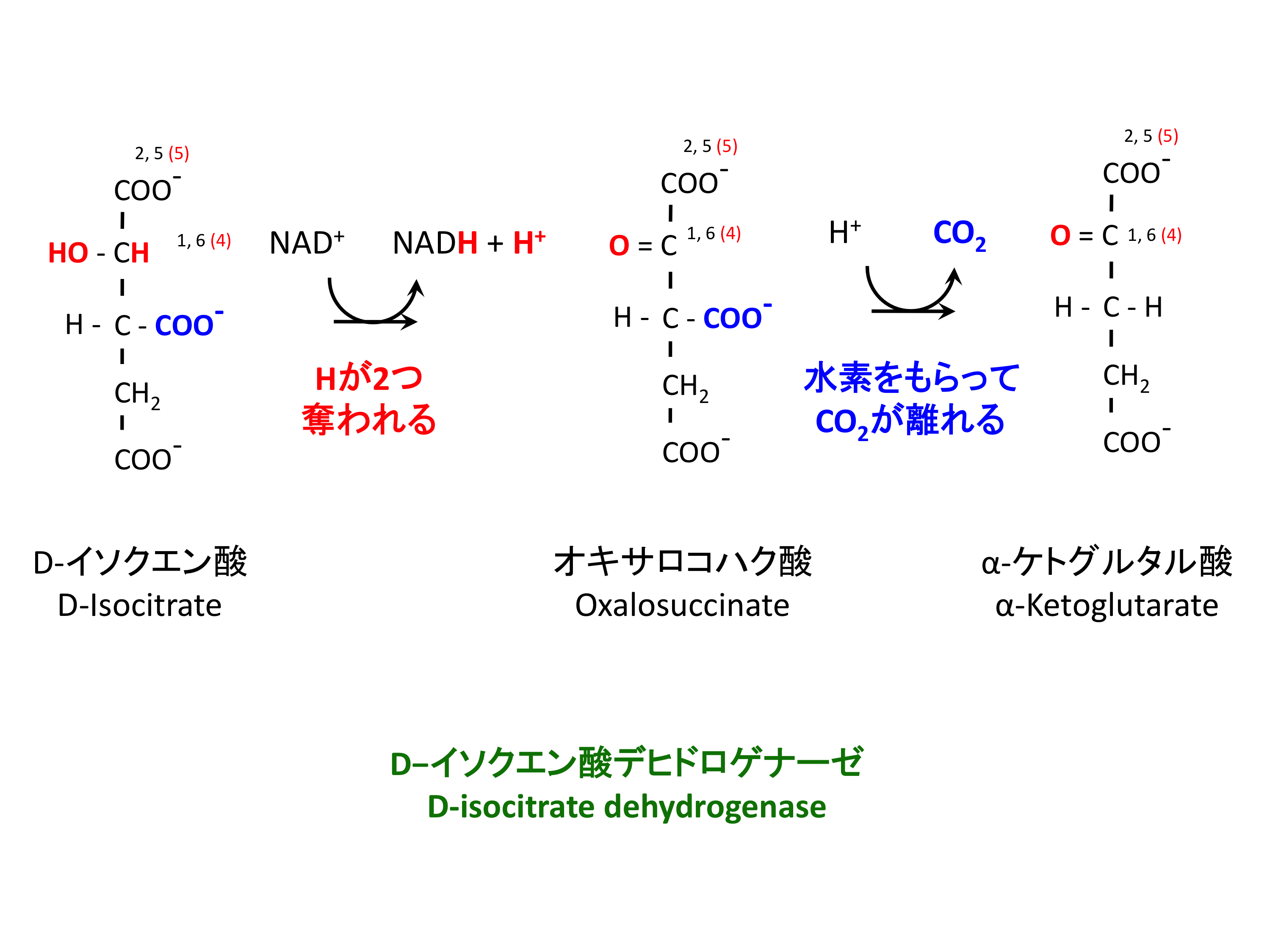
α-ケトグルタル酸は、2-オキソグルタル酸 2-oxoglutarate とも呼ばれる。神経細胞 neuron での グルタミン酸 合成の基質としても重要。
D-isocitrate dehydrogenase は ADP の結合で活性化される (1)。基質との親和性が増大する。ATP および NADH で阻害される。
この酵素が阻害されると、細胞内には D-isocitrate および citrate が蓄積する。両者の変換はきわめて可逆的であるためである (1)。Citrate は細胞質に輸送され、解糖系の律速酵素 PFK を阻害する。つまり、この酵素が律速段階になっているということは、TCA 回路から解糖系へ「TCA 回路が渋滞している」というシグナルを伝える feedback の意味がある。
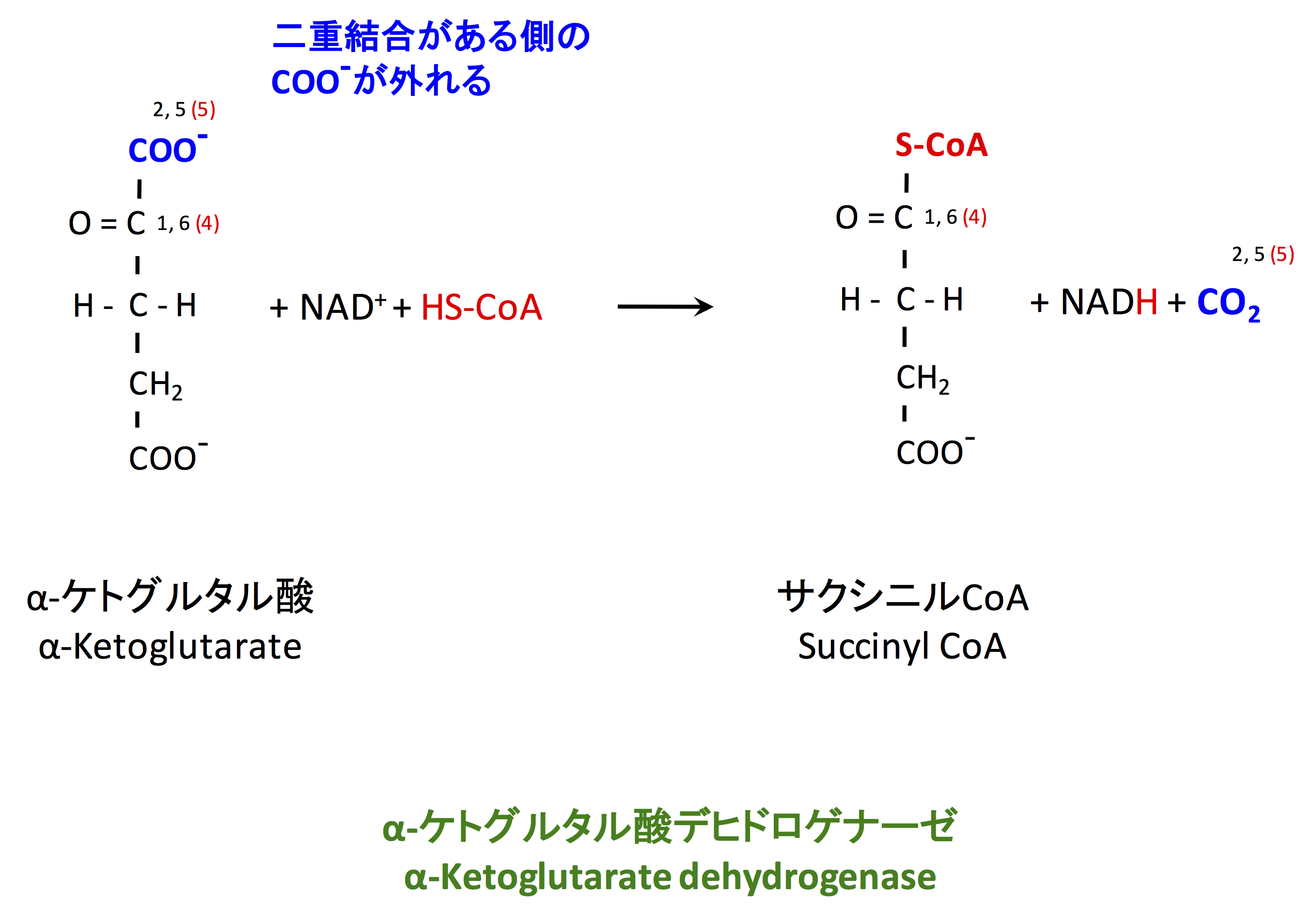
4. 脱炭酸とサクシニル CoA 合成
2 番目の酸化反応は、サクシニル CoA の生成である。この反応を触媒する α-ケトグルタル酸デヒドロゲナーゼは 3 つの酵素の複合体で、ピルビン酸デヒドロゲナーゼに相同である。この酵素は、反応生成物のサクシニル CoA および NADH で阻害される (1)。
実際にこの反応はピルビン酸の酸化・アセチル CoA の生成によく似ており、こちらの図には書いていないが最初に脱炭酸が起こる。
α-ケトグルタル酸 + CoA + NAD+ → サクシニルCoA + CO2 + NADH
ピルビン酸 + CoA + NAD+ → アセチルCoA + CO2 + NADH
広告
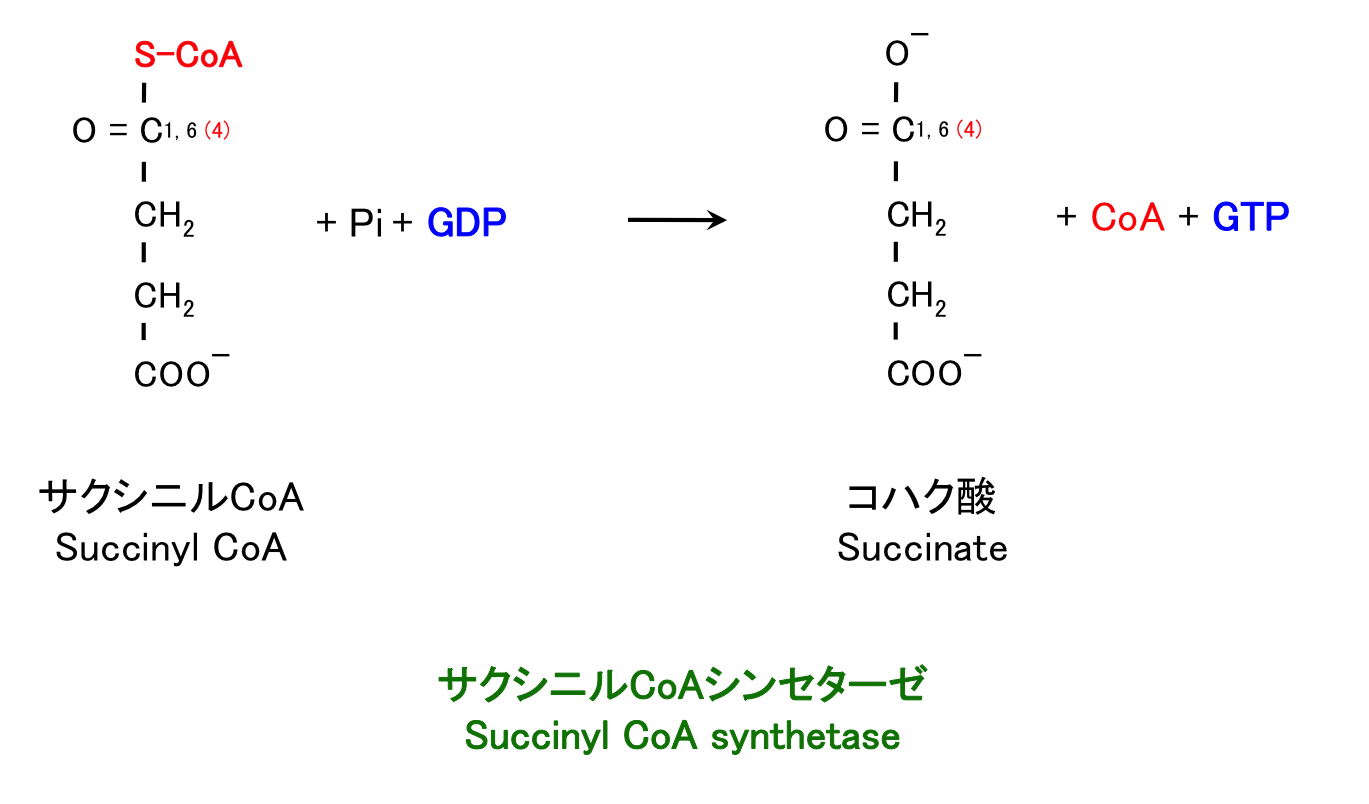
5. サクシニルCoA酸化, GTP 合成
前の反応でできたサクシニル CoA は高エネルギー中間体であり、加水分解される際に放出されるエネルギーは ATP のそれに匹敵する (1)。したがってすぐに CoA が脱離し、そのエネルギーは GDP に受け渡される。
TCA 回路で唯一、直接的に高エネルギーリン酸結合を作る反応である (1)。種によっては GDP でなく ADP を用い、ATP を産生することもある。哺乳類 TCA 回路のこのステップで、ATP でなく GTP が作られることの意義は私にはまだわからない。
GTP はそのままシグナル分子として使われるほか、以下の反応によって ATP を産生する場合もある。
GTP + ATP → GDP + ATP
6. コハク酸の酸化, FADH2 合成
第 3 の酸化反応は、FAD から FADH2 を生成する反応である。ここから続く一連の酸化、水和、酸化という反応は、メチル基 CH3 をカルボキシル基 C=O に変換するときによく使われる反応で、β 酸化 の際にも似たような反応が起こる。
コハク酸 + FAD → フマル酸 + FADH2
この反応では、NAD+ でなく FAD がプロトンおよび電子の受容体として使われる (1)。この反応の自由エネルギーが、NAD+ を還元するのに不十分なためである。このように
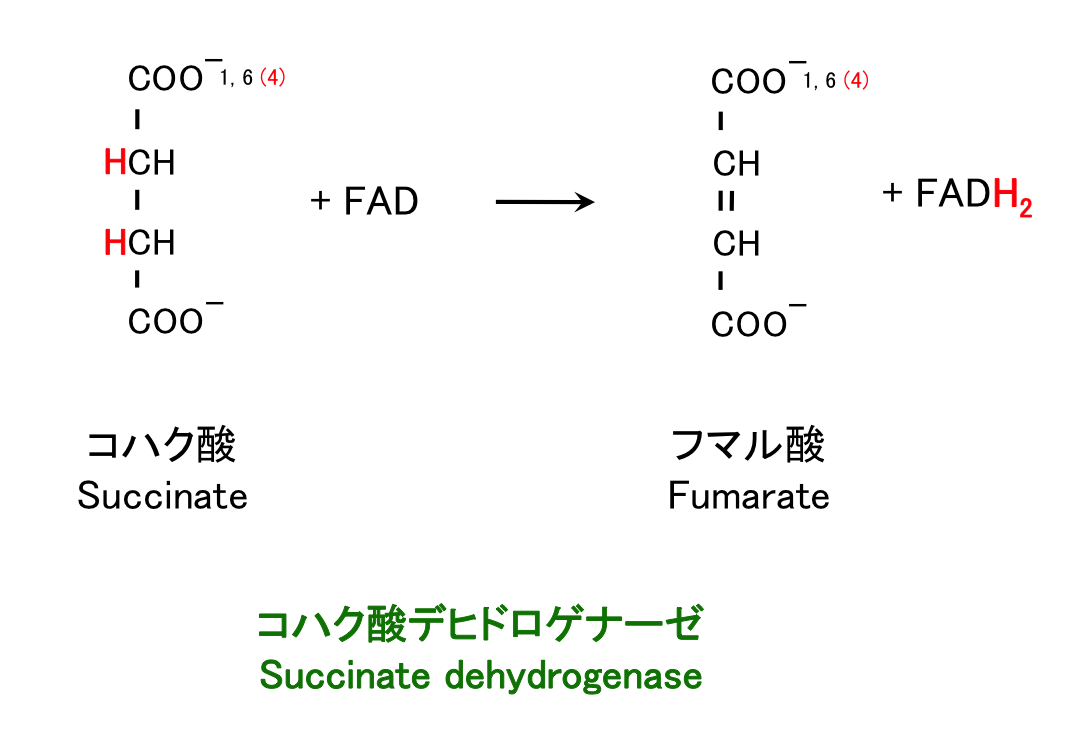
コハク酸デヒドロゲナーゼは、他の TCA 回路の酵素と異なり、ミトコンドリア内膜に埋め込まれており、膜の電子伝達系に直接 FADH2 を渡す (1)。そのため、ページ上の日本語 TCA 回路全体像では、FAD でなく Q が補酵素のように書かれている。
|
参考: しかし、FAD に移った 2 つの水素に着目すると、それ以外の部分の酸化数は反応前後で増えているのである。水素の酸化数は常に +1 なので、コハク酸の −2 は「奪われる水素の +2」と「それ以外の部分の −4」の和である。 一方、フマル酸は水素が奪われたあとなので、全体の酸化数 = それ以外の部分の酸化数 = −2 が成り立つ。 「それ以外の部分」の酸化数は、反応の前後で −4 から −2 へと増加している。つまり、その部分に関しては酸化数が増える酸化反応であると言える。 |
7. フマル酸の水和
フマル酸 fumarate [fjuːməreit] からリンゴ酸 malate [meileit] が合成される。
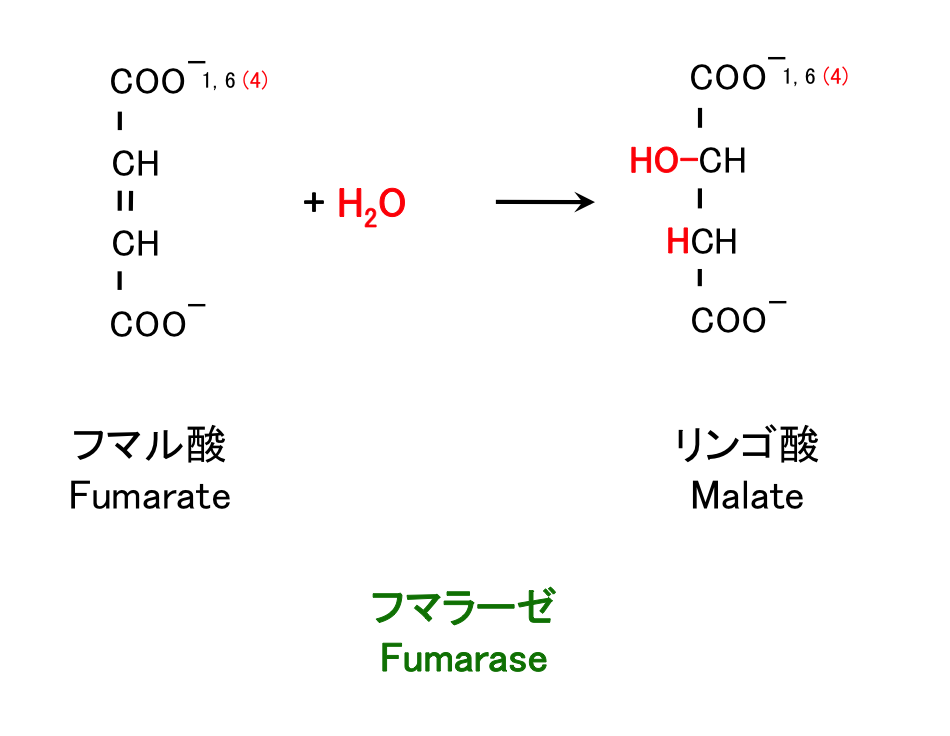
8. リンゴ酸の酸化
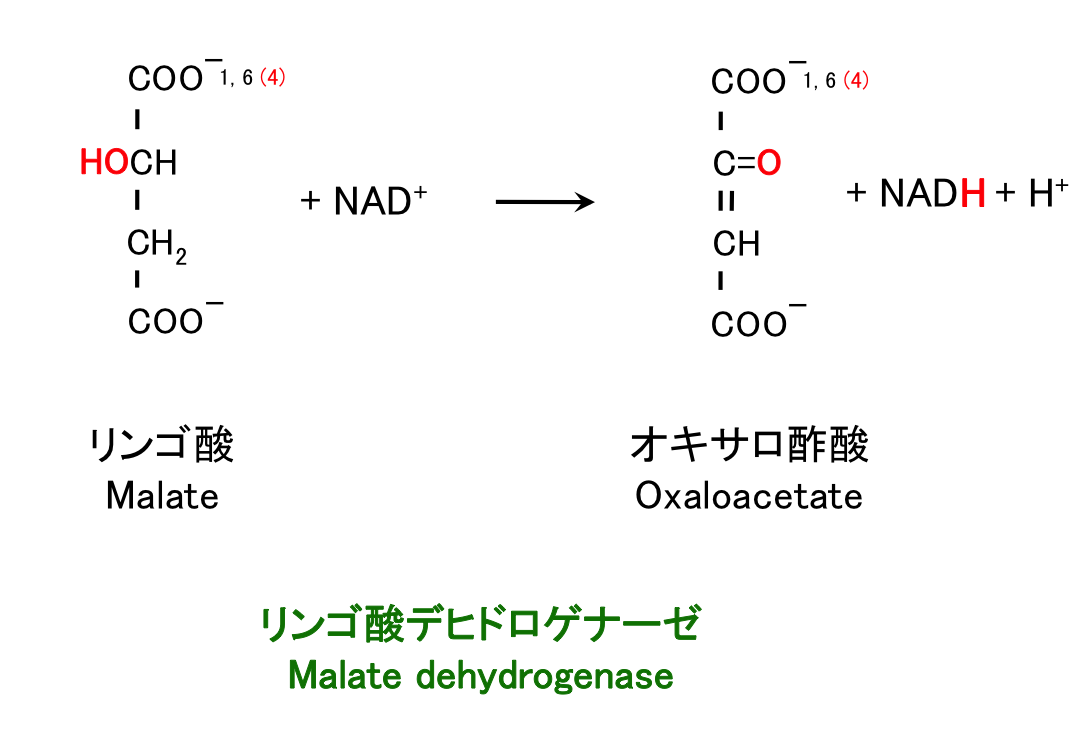
反応 6 – 8 では、生化学反応の典型的なパターンをみることができる。すなわち
- 水素を取り去って C=C 二重結合を作る。
- これを水和して C-OH と CH にする。
- さらに脱水素し C=O にする。
という一連の手順である。脂肪酸の β 酸化で脂肪酸からアセチル CoA を切り取る過程で、この方法がまさに使われている。
TCA 回路の覚え方
お: オキサロ酢酸
く: クエン酸
い: イソクエン酸
あ: α-ケトグルタル酸
さ: サクシニルCoA
こ: コハク酸
ふ: フマル酸
りん: L-リンゴ酸
広告
References
Berg et al. 2006a. (Book) . Biochemistry, 6th edition.
|
Berg, Tymoczko, Stryer の編集による生化学の教科書。 巻末の index 以外で約 1000 ページ。 正統派の教科書という感じで、基礎的な知識がややトップダウン的に網羅されている。その反面、個々の現象や分子に対して生理的な意義があまり述べられておらず、構造に偏っていて化学的要素が強い。この点、イラストレイテッド ハーパー・生化学 30版 英語圏ならば学部教育向けにはややレベルが高い印象。しかし、基本を外さずに専門分野以外のことを |
Jucker et al. 1997a. 13C and 31P NMR studies on the effects of increased plasma free fatty acids on intramuscular glucose metabolism in the awake rat. J Biol Chem 272, 10464-10473.Boumezbeur et al. 2010a . Alterted brain mitochondrial metabolism in healthy aging as assessed by in vivo magnetic resonance specgtrometry. J Cereb Blood Flow Metab 30, 211-221.de Graaf et al. 2004a (Review). In vivo 1H-[13C]-NMR spectrometry of cerebral metabolism. NMR Biomed 16, 339-357.Rothman et al. 1992a. 1H-[13C] NMR measurement of [4-13C]glutamate turnover in human brain. Proc Natl Acad Sci USA 89, 9603-9606.de Graaf et al. 2004b. Regional glucose metabolism and glutamatergic neurotransmission in rat brain in vivo. Proc Natl Acad Sci USA 101, 12700-12705.de Graaf et al. 2003b. Detection of [1,6-13C2]-glucose metabolism in rat brain by in vivo 1H-[13C]-NMR spectroscopy. Magn Res Med 49, 37-46.Chowdhury et al. 2007a. Glutamatergic and GABAergic neurotransmitter cycling and energy metabolism in rat cerebral cortex during postnatal development. J Cereb Blood Flow Metab 27, 1895-1907.Serres et al. 2007a. Brain pyruvate recycling and peripheral metabolism: an NMR analysis ex vivo of acetate and glucose metabolism in rat. J Neurochem 101, 1428-1440.Lardon et al. 2005a. 1H-NMR study of the metabolome of an exceptionally anoxia tolerant vertebrate, the crucian carp (Carassius carassius). Metabolomics 9, 311-323.- Amazon link: 細胞の分子生物学
: 第 6 版を紹介しています。
コメント欄
サーバー移転のため、コメント欄は一時閉鎖中です。サイドバーから「管理人への質問」へどうぞ。