脂肪分解に関わる酵素 HSL: 構造、機能、分子進化など
UBC/protein_gene/h/hsl
このページの最終更新日: 2026/04/09- 概要: HSL とは
- HSL の構造
- HSL 活性の制御
- HSL ノックアウトマウス
広告
概要: HSL とは
脂肪細胞 adipocyte は、油滴 lipid droplet の中に貯めている トリアシルグリセロール を必要に応じて ジアシルグリセロール、ついで モノアシルグリセロール に加水分解し、取り出した 脂肪酸 を血管へ放出する。
この過程を
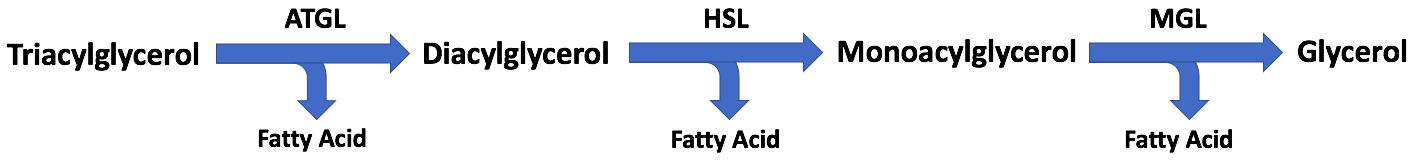
Lipolysis により生じた脂肪酸は、遊離脂肪酸 free fatty acid または非エステル化脂肪酸 (NEFA; non-esterified fatty acid) と呼ばれる。筋肉などに運ばれ、β酸化 などを経てエネルギー源となる。
ホルモン感受性リパーゼ (HSL; hormone-sensitive lipase, EC 3.1.1.3) DAG 分解の律速酵素 で、
HSL のほか、TAG を主に分解する ATGL も重要な酵素である。
> HSL の酵素活性は広い。TAG : DAG : MAG に対する活性は 1 : 10 : 5 である (4)。
- Acylglycerol のほか cholesteryl ester, retinyl ester, steroid ester も分解する (3)。
- Cholesteryl ester に対する分解活性は TAG の約 2 倍で、生理的にどのように働いているのか興味深い (3)。
リパーゼは、至適 pH によって酸性リパーゼおよび中性リパーゼ neutral lipase に分類されることがある。この分類では、HSL は中性リパーゼに属する。
HSL の構造
HSL 遺伝子
哺乳類では、promoter の使い分けによって 3 つの isoform がある。サイズは 84 - 130 kDa と多様 (3)。精巣特異的な HSLtes があり、cholesteryl ester hydrolase として精子形成に必要 (7)。多くの知見は脂肪組織を使った実験から得られており、言及がない場合は HSLadi についてのものと考えられる。
バクテリアも HSL に相同と考えられる分子をもっており、HSL の起源は非常に古いと考えられている。ただし、構造の一部が似ているだけで、その機能には大きな違いがありそうである。
- 哺乳類 HSL は、構造的に bacteria の type B carboxyesterase に似る (5)。
- Bactarial HSL は一般に 34-40 kDa と小さく、哺乳類 HSL の C-terminal region に似る (9I)。
- BFAE from Bacillus subtilis, lipase 2 from Moraxella TA 114などが HSL family protein (9I)。
- Bactarial HSL は水溶性の短鎖脂肪酸は分解するが、エマルジョン中の長鎖脂肪酸はできない (9R)。
- このことから non-lipolytic carboxyestelase と位置づけられる。ヒト HSLは lipolytic である (9R)。
また、HSL は 他のリパーゼ (pancreasic lipase, lipoprotein lipase など)と相同性を示さない (14)。Exon 6 と他のリパーゼの相同性が著しく低く、ここがどこからか飛んできた mosaic protein かも。Exon 6 は phase I (コドンの一番目の塩基で切れる)の構造があり、これも mosaic protein の特徴。
HSLtes は、different promoter usage で5'側に特異的エクソンをもつ (7I)。5' 側の isoform-specific exon が翻訳領域をコードするのは珍しい。大体は非翻訳領域。
哺乳類の脂肪組織、adrenal gland、卵巣では、転写は複数の exon (A, B, C, D, or exon 1 ) から始まる (4)。Exons B, C, D は非翻訳領域なので、どこから転写されるかはアミノ酸配列に影響しない。Exon A は 43 アミノ酸を HSL の N 末端側に付加する。哺乳類の精巣では、2つの組織特異的エクソン (T1, T2)がある (4)。Exon T1 は、300 アミノ酸を N 末端側に付加する。T2 は非翻訳領域をコードする。
タンパク質の構造
- N-terminal domainはタンパク質相互作用 or 基質結合、C-terminal domainは活性ドメイン (4)。
- N-terminal domainは他のタンパク質と相同性を示さない。Cはbactarial HSLsに似る (9I)。
- Rat N-terminal domain (AA1-300 ) は、ダイマー化およびFABP4との結合に関与 (4)。
- Rat C-terminal domain は Ser-423, Asp-703, His-733から成る catalytic triad を含む (4)。
- Rat regulatory module (AA521-669 ) はloop regionで、既知の全てのリン酸化サイトを含む (4)。
- Rat putative lipid binding domain は AA658 あたりにある (4)。
Rat HSL のリン酸化サイト
Ser563: PKA (3,4 ), glycogen synthase kinase-4 (4 )
> site 1, regulatory siteとも呼ばれる (4)。
> 563のみ、または563と565をAlaに置換しても、活性は失われない (4)。
> protein phosphatase 2A (PP2A ) とPP2Cに脱リン酸化される (4)。
Ser565: AMPK (11 ), Ca2+/calmodulin-dependent protein kinase II (4 )
> Site 2, basal siteとも呼ばれる (4)。ここのリン酸化は活性を負に制御するかもしれない。
> かつては活性に影響なしとされていたが、ここに変異を入れると活性がわずかに上昇する。
> AMPKによってリン酸化されると、lipolysis が阻害される (11。
> 主にPP2Aに脱リン酸化されるが、PP1およびPP2Cも重要かもしれない (4 )。
Ser600: Erk (3,4 )
Ser659: PKA (3, 4 )
> Ser660とともに新しく発見されたリン酸化サイト (4)。活性に必要。変異を入れると活性がなくなる。
Ser660: PKA (3, 4 )
広告
「あとがき」で当サイトを参考にしたと書いてくれているラノベです。Kindle Unlimited で読めました。ストーリーと文章が良く、面白かったです。
HSL 活性の制御
転写制御
ヒツジ HSL の 5' 上流域 (転写開始点から 1244 bp)にあった転写因子結合サイト (12)。 Stimulating protein 1 (Sp1 ), CCAAT-box Binding Factors (CBFs ), Activator Protein 2 (AP2 ) and Glucocorticoid Receptor (GR ), as well as other cis-acting regions denominated as Insulin Response Element (IRE ), Glucose Response Element (GRE ), Fat Specific Element (FSE ) and cAMP Response Element (CRE ).
cAMP による制御
> 3T3-F442A および BFC-1 脂肪細胞では、cAMP または PMA 処理で HSL の mRNA 量が低下する (13)。 : PMA (phorbol 12-myristate 13-acetate ) は、PKC の活性化剤である。 : cAMP-PKA 経路、PKC 経路による HSL の活性化を考えると意外な結果。フィードバック? : 短期の飢餓では、HSL の mRNA 量が低下するという報告も引用されている (13D)。
ホルモンによる制御
グルカゴン glucagon および カテコールアミン catecholamine によって活性化される。PKA シグナルを介した作用であることがよく知られている。 > 成長ホルモン growth hormone によって、ニジマス肝臓の HSL がリン酸化される (16)。 : 哺乳類でも示されているのか? : PKC, ERK を介する経路に依存することが阻害剤を使って示されている。
HSL ノックアウトマウス
> KO miceの体重およびWAT重量は正常 (1D)。WATは小さいという報告も (8)。
> HSL-deficient mice are lean, and they effectively mobilize FFAs from TG (2I ).
> KO miceでも basal lipolysis は起こる。これはHSLの他にも lipase があることを示唆する (3)。
: KOではDAGが蓄積するので、HSL以外の lipase はTAG lipaseであることが予想される (3)。
: のちに、TAG の分解活性が高い adipose tissue triglyceride lipase (ATGL) が発見されている。
> DAG 分解活性は、wtの1/20から1/30程度 (1)。
高脂肪食を与えた場合
> 高脂肪食で一般に体重は増えるが、HSL-/-は増え具合がwtよりも穏やか。BATでの脂質燃焼が活発 (8)。
> normal chowおよびhigh-fat dietを与えたHSL-/-は、白色脂肪組織がwtよりも小さくなる (8)。
> HSLはWATでPPARgのリガンドを作っており、KO miceでは脂肪細胞の分化が抑制されるためと考えている。
> high-fat diet を与えた KO mice の BAT では脂質の燃焼が wt より活発。コレステロールの蓄積が原因か? (8)
その他未整理
> KO miceでは血中 NEFA 量が減り、筋肉や心臓でのインスリン感受性が増大する (5)。
> HSL-/- mice は血中 NEFA 量がnormal chowでもhigh-fat dietでも低い (8)。
> HSL-/- mice は不妊である (7I)。
> Drosophilaの油滴タンパク質のリン酸化にも cAMP, PKA が関与。哺乳類lipolysisと似た機構がある (10)。
- リン酸化で lipid droplet の近くへ輸送される (3)。
- PKA, perilipin と共同で脂肪分解をするが、これが adipocytes 以外で保存されているかどうかは不明 (3)。
- 実際に、ペリリピンは脂肪細胞もしくは steroidogenic tissueでのみ発現している (3)。つまり、他の組織での lipolysis では HSL とペリリピンの共同作業が存在しないと思われる。
広告
References
- Haemmerle 2002. JBC, 277, 4806-4815.
- Haemmerle 2006. Science, 312, 734-737.
- Holm 2003 (Review). Biochem Soc Transac, 31, 1120-1124.
Lampidonis et al. 2011aR (Review). The resurgence of Hormone-Sensitive Lipase (HSL) in mammalian lipolysis. Gene 477, 1-11.- Laurell 2000. Protein Engineering, 13, 711-717.
- Park 2005. Am J Physiol Endocrinol Metab, 289, E30-E39.
- Vallet-Erdtmann 2004. JBC, 279, 42875-42880.
- Harada 2003a. Am J Physiol Endocrinol Metab, 285, E1182-E1195.
- Chahinian 2005a. BBA, 1738, 29-36.
Schlegel 2007a (Review). Lessons from "lower" organisms: what worms, flies, and zebrafish can teach us about human energy metabolism. PLoS Genet 3, e199.Daval et al. 2005a. Anti-lipolytic action of AMP-activated protein kinase in rodent adipocytes. J Biol Chem 280, 25250-25257.Lampidonis et al. 2008a. Cloning and functional characterization of the 5' regulatory region of ovine hormone sensitive lipase (HSL) gene. Gene 427, 65-79.Plee-Gautier et al. 1996a. Inhibition of hormone-sensetive lipase gene expression by cAMP and phorbol esters in 3T3-F442A and BFC-1 adipocytes. Biochem J 318, 1057-1063.- Langin et al. 1993a. Gene organization and primary structure of human hormone-sensitive lipase: possible significance of a sequence homology with a lipase of Moraxella TA144, an antarctic bacterium. PNAS, 90, 4897-4901.
- Molecular Biology of the Cell, 5th edition.
- By MHarberson - Own work, CC BY-SA 4.0, Link
コメント欄
サーバー移転のため、コメント欄は一時閉鎖中です。サイドバーから「管理人への質問」へどうぞ。