グルタミン酸の構造と機能: 神経伝達、旨味、生合成など
- 概要: グルタミン酸とは
- グルタミン酸の生合成
- グルタミン酸の分解
広告
概要: グルタミン酸とは
グルタミン酸 glutaminc acid は図のような構造をもつアミノ酸である。略号は E または Glu。イオンの状態では glutamate という。

pH 7 前後の水溶液中では、2 つの COOH 基が COO- に、NH2 は NH3+ になっている。つまり、このアミノ酸の側鎖から H+ が放出されているということで、酸・塩基の定義 よりグルタミン酸は
また、側鎖の COOH 基が電離していることから、グルタミン酸は負に荷電している。なお、側鎖の COOH 基が CO-NH2 になっているのがグルタミン Gln である。
- pK1 (COOH) = 2.1
- pK2 (NH3+) = 9.5
- pK3 (側鎖) = 4.1
グルタミン酸は、1866 年にドイツの科学者 Karl Ritthausen によって単離された。グルテン (小麦中のタンパク質の総称) の加水分解物から発見された (10) ため、この名がつけられている。
グルタミン酸には以下のような性質がある。
タンパク質構成アミノ酸として
- 負電荷をもっており、タンパク質の表面に位置して水溶性を付与する (9)。
- タンパク質をポリグルタミン酸テールとして修飾することがある (9)。たとえば tubulin は poly-Glu 修飾を受け、これによって他のタンパク質との結合性に影響が出る。
- ビタミン K 依存的なカルボキシル化を受けて、カルシウム結合性が増大する (9)。
遊離アミノ酸として
全て Ref 9 より。
- 摂取した Glu は消化の過程で 70 - 90% 以上が分解されてしまう。体内の遊離 Glu は生合成されたもの。
- システイン Cys、グリシン Gly とともに、トリペプチドであるグルタチオン glutathione の材料となる。
興奮性の神経伝達物質 であるほか、抑制性の神経伝達物質 GABA を合成する材料になる。詳細は グルタミン のページにグルタミン - グルタミン酸サイクル の説明があるので、そちらを参照のこと。- 膵臓 β-細胞からのインスリン insulin 分泌を制御するシグナル分子としてはたらく。
- L 体のモノナトリウム塩 sodium glutamate は旨味を呈し、味の素の主成分。IMP との相乗効果がある。D 体は美味でない (1)。
広告
「あとがき」で当サイトを参考にしたと書いてくれているラノベです。Kindle Unlimited で読めました。ストーリーと文章が良く、面白かったです。
グルタミン酸の生合成
Glu はタンパク質を構成するだけでなく、遊離アミノ酸としても重要な生理機能をもつが、ヒトやラットでは餌に含まれる Glu は腸でほぼ全て分解されてしまう (9)。したがって、
グルタミン酸は、TCA 回路 の α-ケトグルタル酸 α-ketoglutarate を基質として合成される。さらに以下の 2 つの反応にわけることができる。これは、以下の図にみるように両者が構造的に似ていて、アミノ基の付加を付加するだけで良いためである。
アンモニウムイオンを使う場合
グルタミン酸デヒドロゲナーゼに触媒される反応である (6)。デヒドロゲナーゼは脱水素酵素なので、この反応は逆反応である。
NH4+ + α-KG → Glu + H2O
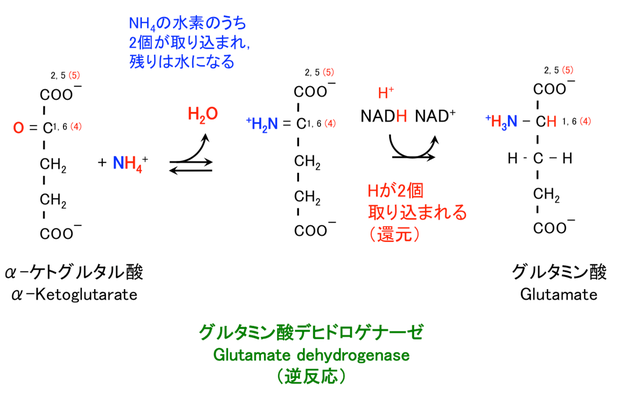
NADH のほか、NADPH が使われることもある。NADH, NADPH の両方を使うことができる珍しい生化学反応である。
炭素番号は α-KG が グルコース 由来である場合の、もとのグルコースの炭素番号である。
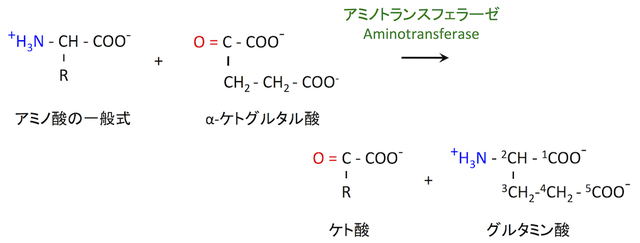
他のアミノ酸を使う場合
アンモニア でなく、他のアミノ酸からアミノ基を転移することでグルタミン酸を合成する経路もある。アミノトランスフェラーゼによって触媒される。
グルタミン酸の合成というよりは、他のアミノ酸を分解する過程で起こる反応と捉えた方がよい。
グルタミン酸の分解
グルタミン酸デヒドロゲナーゼ glutamate dehydrogenase の作用でアミノ基が取り去られ、α-KG になる (6)。上記の生合成の逆反応である。α-KG は TCA 回路に取り込まれる。
- 主に肝臓で起こる。この反応は常に平衡に近い状態にあり、単純に基質の量で合成・分解のどちらに流れるかが決まる (6)。
- グルタミン酸デヒドロゲナーゼは ミトコンドリア に局在する。有毒な NH4+ を隔離するためと考えられており、実際に 尿素回路 もミトコンドリアにある (6)。
広告
References
- Amazon link: 岩波 理化学辞典 第5版
: 使っているのは 4 版ですが 5 版を紹介しています。
- Schmitz et al. 2011a. Cellular changes underlying hypoxia-induced delay of white matter development. J Neurosci, 31, 4327-4344.
- Fitzpatrick et al. 1990a. The flux glucose to glutamate in the rat brain in vivo as determined by 1H-observed, 13C-edited NMR spectrometry. J Cereb Blood Flow Metab, 10, 170-179.
- de Graaf et al. 2011a (Review). State of the art direct 13C and indirect 1H-[13C] NMR spectrometry in vivo. A practical guide. NMR Biomed 24, 958-972.
- Govindaraju et al. 2000a. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed 13, 129-153.
- Berg et al. Biochemistry: 使っているのは 6 版ですが 7 版を紹介しています。
- de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectrometry of cerebral metabolism. NMR Biomed 16, 339-357.
- de Graaf et al. 2004b. Regional glucose metabolism and glutamatergic neurotransmission in rat brain in vivo. Proc Natl Acad Sci USA 101, 12700-12705.
- Brosnan and Brosnan 2013a (Review). Glutamine: a truly functional amino acid. Amino Acids 45, 413-418. See blog.
- Amazon link: 脳単―ギリシャ語・ラテン語 (語源から覚える解剖学英単語集 (脳・神経編)).
- Chouwdhury et al. 2008a. Chronic riluzole treatment increases glucose metabolism in rat prefrontal cortex and hippocampus. J Cereb Blood Flow Metab 28, 1892-1897.
- Patel et al. 2001a. Glutamine is the major precursor for GABA synthesis in rat cortex in vivo following acute GABA-transaminase inhibition. Brain Res 919, 207-220.
- Patel et al. 2005a. The contribution of GABA to glutamate/glutamine cycling and energy metabolism in the rat cortex in vivo. PNAS 102, 5588-5593.
コメント欄
サーバー移転のため、コメント欄は一時閉鎖中です。サイドバーから「管理人への質問」へどうぞ。